




서론
우유의 살균은 우유 안전을 향상시키는 효과적인 방법이지만, 최근까지 미국에서 살균되지 않은 우유의 섭취로 공공 보건 문제가 발생하고 있다는 사실에 조금은 놀라지 않을 수 없다. 살균되지 않은 우유의 섭취는 Campylobacter spp., Shiga toxin-producing Escherichia coli, Salmonella enterica serotype Typhimurium과 같은 여러 병원균에 의한 심각한 질병과 관련이 있다.
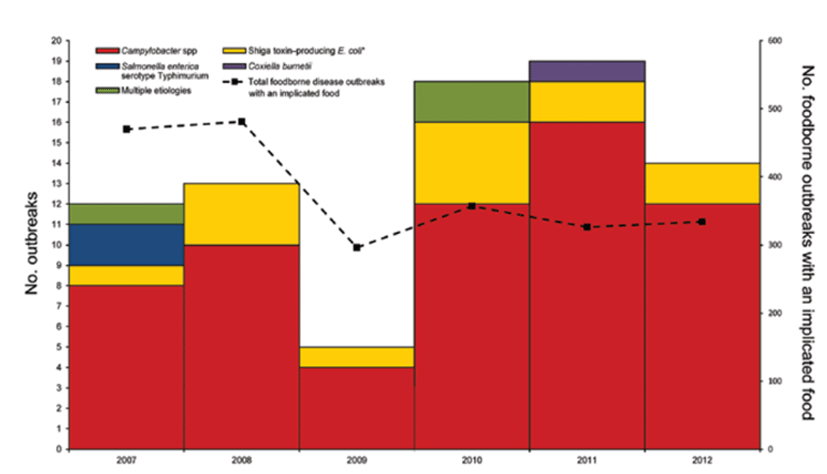
사실, 유럽과 북미에서 우유를 살균해야 한다는 거세 대한 법제화가 이루어진 이후에, 유제품에 사용되는 원유를 반드시 살균해야 한다는 규정은 미국의 미시간 주에서 1948년에 처음 제정되었다. 1987년 미국 식품의약국(FDA)은 살균하지 않은 우유의 전국적 판매(Interstate sale)를 금지시켰다. 그렇지만 State에서의 판매를 규제하는 법률은 각 주마다 다른데, 비살균 우유의 전국판매에 대한 규제는 완전 금지에서 농장이나 소매판매점(retail outlet)의 판매 허용까지 다양하다. 비살균 우유의 판매가 불법인 주에서도 cow-share나 herd-share agreement(협정)을 통해 구할 수 있는데, cow-share agreement는 구매자가 생산된 우유의 비율에 대한 대가로 구매자가 위탁사육료를 목부에게 지불하는 제도이다. 비살균 우유 음용이 보건상의 문제가 있음에도 불구하고, 비살균 우유에 대한 수요가 증가하고 있는 점은 흥미로운 일이다. 미국에서 비살균 우유로 인한 발병 건수는 2007~2009년 30건에서 2010~2012년 51건으로 증가했으며, Campylobacter spp.로 인한 발병이 가장 높았다(77%).
“우유의 열처리가 우유품질과 영양가에 미치는 영향”의 세 번째 장은 “우유 열처리에 의한 병원균 사멸효과”라는 제목으로 중앙대학교 윤영호 교수가 작성하였다. 원유 및 우유에 존재하는 병원성세균에 의하여 발생되는 질병에 대한 내용과 열처리 효과에 대한 연구결과들이 자세하게 제시되어 있어, 후학들이 공부하기에 충분한 가치가 있다고 판단된다. 그러나 본문 내용 중에서 학문 후속세대들이 이해하기 어려웠던 것은 가감하여 재작성하였으며, 원전에 인용문헌이 자세하게 기술되어 있지 않았기 때문에, 논문 재작성시 일일이 찾아 학술지 인용문헌 형식으로 바꾸었지만, 찾을 수 없었던 논문은 부득이하게 삭제했음을 밝혀둔다.
본론
우유를 매개체로 하여 병원균이 전염되는 예가 1900년대 이후 여러 가지 경로를 통하여 나타나는 것이 확인되었으며, 이러한 우유매개성 질병과 병원균의 전염과정에 관한 많은 연구결과가 보고되어 있다(Bryan, 1983).
우유매개성 질병의 발생은 대부분의 경우, 열처리되지 않은 생유음용과 연관하여 나타났다. 이는 대부분의 병원성 미생물은 저온살균 및 열처리 과정에서 살멸된다는 사실을 확인해 주는 것이다.
오늘날에도 우유 매개성 질병과 그 병원체로 중요시 되는 것은 Salmonellosis, Campylobacteriosis, Staphylococus 중독증, Brucellosis, Yersiniosis 등이 있다.
이들을 중심으로하여 과거에 문제시되었던 우유매개성 질병의 병원균이 열처리에 대한 저항성과 저온살균의 가열 살멸 효과에 관하여 고찰하기로 한다.
우유매개성 질병과 병원체
우유매개성 질병은 그 발생빈도를 기준하여, 1) 근래에 중요시되는 질병과 그 병원체, 2) 과거에 관심이 집중되었던 우유매개성 질병과 그 병원체, 3) 우유매개는 희소하게 나타나는 질병과 병원체로 구분될 수 있다.
1900년대 초기로부터 1945년 기간 중에는 주로 생유음용과 연관되어 질병이 발생되었으나, 그 이후 대부분의 우유는 살균되었으므로 발생률은 급격히 감소하였다.
우유의 저온살균은 1900년대 초기에 공중보건학적으로 여러 문헌에 보고되었지만, 널리 보급되지는 않았다. 미국의 경우, 미국 보건성에 의해 발표된 우유처리규정은 1924년에 제정 발표되었고, 그 이후부터 저온살균이 국가적 차원에서 강조되었다. 1950년대 이후에는 생유와 연관된 질병 발생 건수가 극히 소수이지만 이 기간에도 발생보고는 있으며, 개인적 기호에 따라 열처리하지 않은 우유음용을 선호하기 때문인 것으로 판단된다.
Table 1에는 10년을 주기로 하여 미국에 있어서 우유매개성 질병 발생빈도를 나타내었으며, 여기에 제시된 뚜렷한 경향은 1900년부터 1940년까지는 장티프스가 우유매개성 주요 질병이었으나, 그 후 20년간 그 발생률은 급격한 감소를 나타내어 1960년 이후에는 장티프스 발생이 우유와 연관하여 발생되었다는 보고는 없었다.
1900년으로부터 1930년 기간에 발생빈도가 두 번째로 높았던 질병은 연쇄상구균 감염증(성홍열)이었으나, 그 이후 이 질병 발생도감소하여 현재에는 거의 발생보고가 없는 상태이며, 디프테리아는 간혹 발생 사례가 보고되었으나 1946년 이후에는 나타나지 않았다.
포도상구균성 식중독증과 살모넬라 감염증 및 병원균을 알지 못하는 질병 등이 근래에는 관심의 대상이 되었는데, 주로 아이스크림, 분유, 치즈 등이 이 질병 발생과 연관된 것을 확인할 수 있었다(Bryan, 1983).
살모넬라균증과 병인체를 알 수 없는 위장염이 1970년대에서 중요한 생유 매개성 질병으로 확인된 바 있으며, Campylobacteriosis도 중요한 질병으로 부상하였다.
1970년대에 우유매개성 질병 발생에 있어 매개체로 작용한 우유 및 유제품의 종류는 Table 2에 제시되었다. 가정에서 제조한 ice cream에는 계란을 첨가하는데 이 기간 중 Salmonella 감염증 발생과 가장 높은 빈도로 연관성이 있는 것이 확인되었다. 치즈와 생유가 빈번하게 질병매개 유제품으로 지적되었고, 일반적으로 우유로 표시된 것은 생유를 지칭하며 저온살균유와 chocolate milk는 부적절한 살균을 했거나 살균 이후 2차 오염되었을 것으로 판단되었다.
우유열처리에 의한 병원균 사멸효과
살모넬라 감염증(Salmonellosis): 살모넬라 감염증이 우유매개성으로 발생하는 경우는 우유를 열처리 또는 끓이지 않고 섭취하는 지역에서 빈번히 발생한다.
그러므로 생유와 특급우유(certified milk)가 매개체로 판명되었고 때에 따라 살균유, 분유, Cheddar cheese도 연관되는 경우가 있다.
Salmonella dlublin, Salmonella newbrunswick, Salmonella geiderberg, Salmonella derby 등이 병원균으로 분리되었다.
우유 중에 이 병원체가 오염되는 경로는 사료 또는 우사 내의 환경으로부터 감염되어 분변으로 배설하게 되며, 소의 피부와 유방이 이 병원체로 오염되어 우유로 이행되어 재오염되는 경우도 있다.
열처리에 의한 Salmonella의 사멸효과는 55℃에서 60분간, 60℃에서 15~20분간 가열할 때 사멸된다. 전지유에서 Salmonella는 62.8℃에서 3.6~5.7초 이내에 사멸하고, 65.6℃에서 1.1~1.8초, 68.3℃에서는 0.28~0.52초 가열에 의해 사멸한다(Read et al., 1968).
Salmonella 균주 중에서 가장 열저항성이 높은 균주는 Salmonella Senftenberg 775 W로서 65.5℃에서 34초, 68℃에서 10초, 71.7℃에서 1.2초, 73.9℃에서 0.55초 가열할 때 사멸한다. 따라서 모든 살모넬라균주가 사멸함에 있어 62.8℃에서 18.4초만 가열하더라도 충분한 살균효과가 있을 것으로 판단된다.
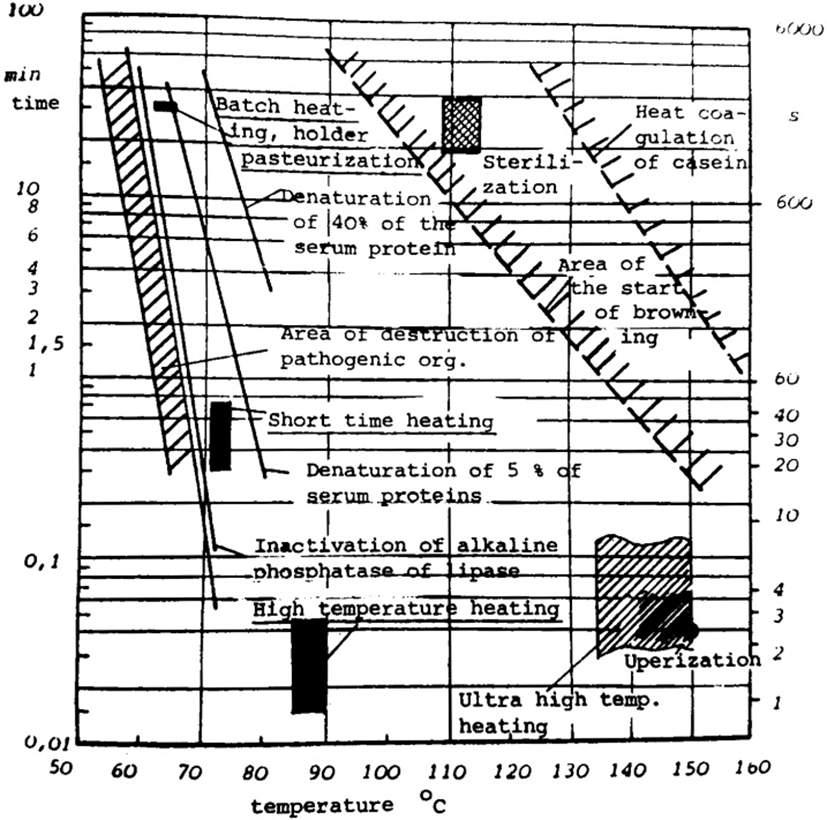
Salmonella sp.를 포함한 여러 종류의 병원성 비포자형성 세균의 가열살멸 조건과 그 내열성은 Fig. 2에 나타내었다(Kessler, 1981). 또한 UHT 열처리과정에서 포자사멸조건과 내열성에 관하여 제시되어 있다.
켐피로박터 감염증(Campylobacteriosis): 이 질병은 미국, 영국, 스코트랜드, 캐나다에서 우유에 의해 매개되어 발생된 보고가 있으며 주로 생유와 특급우유가 매개체로 작용된 것으로 밝혀졌다.
발생된 환자의 분변에서 Campylobacter jejuni가 분리되었고 병원체로 확인되었다. 많은 종류의 포유동물이 C. jejuni에 감염될 수 있으며, 이 병원체가 사람, 소, 면양, 돼지, 닭, 야생조류의 분변에서 분리된 바 있다. 또한 이 병원체는 계육, 해수, 담수 등에서도 분리된 바 있어 이를 통해서 유우 혹은 우유에로 오염 될 수 있다.
Campylobacter jejuni는 56℃에서 5분간 가열하면 사멸되므로 저온살균과정에서 충분히 사멸효과가 나타날 수 있다(Waterman, 1982).
C. jejuni는 가열에 대하여 민감하여 55℃에서 D치는 peptone 중에서 2.25분이고, C. fetus subsp. jejuni의 탈지유 중에서 사멸성향에 관한 실험(Doyleetal, 1981)에서 48℃에서 D치가 7.2~12.8분, 55℃에서 0.74~1.0분이었으며, 우유 중에 오염도가 높을 때 열처리조건을 55℃에서 10분간 가열하면 충분히 살균되는 것으로 확인하였다.
Gill 등(1981)에 의하면 이 병원균이 9×103~2.3×106/mL 수준으로 오염된 생유를 고온단시간살균법(HTST)으로 처리될 때 충분한 살균효과를 얻은 것으로 확인하였고, Christopher 등(1982)은 탈지유 중에서 C. jejuni와 C. intestinalis는 60℃에서 1분 가열에 대하여 생존하지 못하였고, 50℃에서 D value는 C. jejuni 는 1.3~4.5분, C. intestinalis는 1.0~3.7분이라고 보고하였다.
C. fetus subsp. jejuni 5균주를 48℃ 50℃, 53℃ 및 55℃에서 사멸시키는데 소요시간을 측정한 결과는 Table 3에 제시된 바와 같으며, 균주에 따라 55℃에 D value가 0.74~1.0분으로 차이를 내었다(Doyle and Roman, 1981).
| D value (min) at | ||||
|---|---|---|---|---|
| 48℃ | 50℃ | 53℃ | 55℃ | |
| F R I - CF 3 | 12.8 | 5.4 | 1.57 | 0.74 |
| F R I - CF 6 | 7.2 | 4.7 | 1.83 | 1.00 |
| F R I - CF 8 | 7.7 | 3.5 | 1.85 | 0.93 |
| F R I - CF 12 | 12.8 | 4.4 | 1.56 | 1.00 |
| F R I - CF 16 | 11.7 | 5.1 | 1.95 | 0.86 |
포도상구균성 식중독(Staphylococcal intoxication): 착유 이후 바로 냉각하지 않은 생유나 저온살균이 보편화되기 이전의 우유, 산양으로부터 착유된 생유가 포도상구균성 식중독을 일으키는 매체가 된 것으로 확인된 바 있다. 드물게 나타나는 예로서 열처리한 후 재오염되어 발병한 예도 있으며 분무 건조된 분유에 의해서 포도상구균 감염증이 발병예가 있고, 치즈, 아이스크림, 버터도 매개체로 작용한 예가 보고된 바 있다.
Staphylococcus aureus가 대표적인 병원균으로 확인되었고 이 병원균은 유우의 유방과 유관(teat canal)에서도 자주 발견된다. 이는 또한 유방염의 원인균이 되며 우유유제품이 이 병원균에 의해 오염되는 경로는 낙농가공 공장의 종업원들의 손이나 콧물에 의해 오염되며, 이 병원체의 감염에 의한 증상이 나타남에 있어서는 충분한 량의 enterotoxin이 생산되어야 한다.
이 병원균의 열저항성은 낮은 수준이며 100균주를 65℃에 3분간 가열할 때 생존하는 균주는 3균주에 불과하였고 63~64℃에서 30분 가열할 때에는 완전히 사멸한다. Enterotoxin을 생산하는 배양조건은 원유 중에서 23℃에 24시간, 살균유에서는 12시간 배양하면 독소생산이 가능한 것으로 보고한 바 있다(Bonzovic, 1963).
또한 coagulase를 생산하는 Staphylococcus 5균주를 우유 중에서 가열할 때 60℃에서 18분, 63℃에서 5분, 66℃에서 2분, 71.6℃에서 1분 가열하면 사멸하였고, 열저항성과 병원성은 상관관계가 있음을 확인하였다. 우유에서 분리된 Staphylococcus aueus 를 우유에 배양하고 가열처리할 때 60℃에서 D value는 0.87분이고, 염류를 함유하는 배지에서 D value는 0.62분이었으며, 이 병원균은 가열시작 직후로부터 가열손상을 입으며, 온도증가에 따라 가열충격도 증가하며, 70℃에 도달하면 가열손상과 사멸이 병행하는 것으로 확인하였다. 또한 Z value는 9.46℃로 확인하였다(Firstenber 등, 1977).
생유 중에서 enterotoxin을 생산함에 있어서 S. aureus를 103~ 105 /mL 농도로 접종한 후 6~8 시간 이후에는 enterotoxin이 측정 가능한 농도에 이르기까지 생산되었으나, 61℃에서 18초간 저온살균처리된 우유에 같은 수준으로 접종한 경우에는 생균수가 4~6시간이 후에 1.2×106 /mL 수준에 도달하고 독소가 생산되어 열처리된 우유에서 성장이 생유의 경우보다 용이한 것으로 확인되었다.
S. aureus는 enterotoxin을 생산함으로 구토를 수반하는 증상을 보이며 독소가 섭취되면 위장벽을 자극하고, 설사증상도 나타나지만 그 원인은 밝혀지지 않았다. S. aureus가 생산하는 enterotoxin은 분자량 25,000~35,000 dalton의 단순 단백질로서 열에 대한 안정성은 비교적 높다. Enterotoxin B가 A, D보다 열저항성이 높으나, pH에 큰 영향을 받으며, enterotoxin B를 완충용액에서 110℃ 8분 가열 시 불활성화되었으나, beef broth에서 파괴시키는 데에는 110℃에서 60분이 소요되었다. Beef broth 중에서 가열파괴 시간이 연장되는 현상은 단백질의 보호 작용에 기인된 것으로 보인다(Lee, 1977).
브루셀라증(Brucellosis): 열처리하지 않은 산양유로 만든 cheese가 브루셀라증과 연관이 있는 사례가 보고되었으며, 특히, Mexico 지역에서 생산된 산양유 cheese가 문제시되고 있다.
젖소, 유산양, 물소 유 젖을 생산하는 동물이 Brucella의 보균자가 될 수 있으며, 이 병원균은 임신된 암컷의 자궁과 비유 중에 있는 동물의 유선에 집합되어 있어 감염된 동물은 수년간 비유를 통하여 Brucella를 배출하게 된다.
Brucella는 감염된 가축과 접촉하거나, 배설물, 먼지 등에 의해 오염되며, 이들은 생유 중에 여러 시간동안 생존이 가능하고 cheese에서는 수주일간 생존한다.
Brucella abortus에 오염된 원유를 63℃에서 30분간 저온장시간 살균과 HTST(72℃ 15초) 처리한 후 guinea pig에 접종하여 혈청학적 실험과 세균학적 실험을 병행한 결과, 음성으로 확인되어 소비자에게 안전하다는 사실이 확인되었다(Heever et al., 1982).
Foster 등(1953)의 연구에서 Brucellaabortus 2308이 우유에 오염되어 있을 때 142° F에서 23분, 161°F에서 45초간 가열될 때 살멸되었다고 보고하였다.
Yersinia증(Yersiniosis): 생유를 매개체로 하여 맹장염과 증상이 유사한 장염이 발생한 예가 보고되었으며, chocolate milk도 매개체의 역할을 한 예가 보고된 바 있다.
Yersinia enterocolitica에 의하여 장염 증상이 나타나지만 Yersiniapseudotuberculosis에 의하여 패혈증과 설사 및 용혈성 증상이 나타나기도 한다. 동물이 Y. enterocolitica 감염된 경우는 자주 보고되고 있으며 개울물에도 이 병 원균이 오염된 예가 있어 우유도 이러한 감염원에 의하여 오염된 것으로 추정하고 있다.
Y. enterocolitica를 인위적으로 우유에 첨가한 다음 저온살균한 후 시료를 평가한 결과, 100 시료 중 단지 1개의 시료에서만 분리 가능하였으므로 저온살균에 의하여 살균이 효과적으로 이루어진다는 사실이 확인되었다.
장티프스: Salmonella에 의해 발병되는 질환을 그 병상에 따라 4종류로 구분되며, 이 증상은 개별적으로 나타나기도 하나 동시에 나타나는 경우도 있다. 증상이 특징적으로 나타나지 않는 회복기의 증상이 있고 장티프스와 파라티프스의 경우와 같이 고열증상을 수반하는 경우도 있으며, 식중독증상에서 나타나는 장염증상과 패혈증의 증상을 보이는 것 등이다.
장티프스는 우유에 저온살균이 보편화되기 이전 시기에는 우유를 매개체로 하여 빈발했던 질병이며, 그 병원체는 Salmonella Typhi 이다. 장티프스에 이환되었던 사람이나 이 병원균의 carrier인 사람 혹은 가족 중 장티프스 환자를 간호하던 사람이 감염원이 될 수 있고, 보균자들이 젖소의 착유 또는 우유용기를 취급함으로 인하여 우유에 2차 오염된다.
이와 다른 오염원으로 우유용기 및 기구를 세척하는 물을 통하여 오염되는 예도 있으며, 환자의 가정으로부터 환수된 우유병을 통하여 오염될 수도 있다. Salmonella Typhi를 열처리된 우유에 오염시키고, 열내성을 조사한 결과를 보면 전지유에 101 균주의 장티프스균을 접종한 후 71℃에서 19초간, 85℃에서 12초간 가열할 때 100 균주가 사멸하는 효과를 확인할 수 있었던 것으로 보고된 바 있다.
성홍열(Scarlet fever): 우유에 의해 매개된 성홍열 발생보고는 미국에서 있었으며, 저온살균이 보편화되기 이전에는 전 세계적으로 발생된 바 있다. 이 병원균이 우유에 오염되는 경로는 유방염에 이환된 유우의 유방과 유두 등에서 착유 시 원유로 이행하게 된다. 사람이 carrier가 되어 소에 A형 유방염이 발생되는 예가 있으며, 이 경우 환축은 많은 수의 연쇄상구균 (streptococci)을 우유로 방출하게 된다. 이러한 우유를 열처리하지 않고 마시게 되면 발병하게 되는 것이다.
Streptococcus pyogenes가 주요 원인균이며, 이 미생물은 56℃에서 30분간 가열시 살멸되므로 저온살균이 적정하게 이루어질 경우 살균된다.
디프테리아: 저온살균이 보급되기 이전에는 디프테리아 발생이 우유와 연관되어 나타난바 있다. 목장에서 일하는 사람이 Corynebacterium diphtheriae의 carrier이거나 환자일 경우 원유에 2차 오염될 수 있다.
이 병원균은 가열에 의하여 쉽게 사멸되는 미생물로 원유에 오염된 상태에서 58℃에 10분간 열처리로 사멸한다.
우유 중에서 열내성을 측정한 결과를 보면 143°F에서 0.064분, 161°F에서 0.096초이며, chocolate milk 중에서는 143°F에서 0.025분 161°F에서 0.02초로 확인된 바 있다.
결핵(Tuberculosis): 소 결핵균 Mycobacterium bovis는 원유에 의하여 전파될 수 있으며, 인체 내에 침입한 병원균은 후두 및 장에 침투하여 경부 임파절에서 병변을 생성케 한다.
원유를 매개체로하여 Mycobacterium bovis가 사람에게로 전파될 수 있다는 증거가 제시된 바 있으며, 그 전파에 영향을 주는 요인 중에는 유우 중에 감염을 일으키는 조건, 즉 M. bovis가 오염될 수 있는 기회와 원유 중에 존재하는 미생물 수, 원유의 열처리 여부 및 우유 마시는 습성 등이 있다. 많은 수의 결핵균에 감염된 동물의 젖에는 M. bovis가 나타날 수 있으며, 이 병원균은 분변에서 유래하여 젖소의 유방, 엉덩이, 꼬리 등에도 존재할 수 있다. 또한, 감염된 유방이나 호흡기에 존재하는 병원균으로부터 오염될 수도 있다. 동물에서 우유로 오염된 예는 드물고, 열저항성을 보면 저온살균과정에서 M. bovis의 많은 수가 사멸한다.
액상 유제품에서 결핵균의 가열에 의한 사멸 연구(Stemin, 1958)에서 Mycobacterium tuberculosis 2균주를 우유와 ice cream mix에 가하고, 70℃와 82℃에서 각각 열처리하고, 일정한 시간간격으로 시료를 채취하여 guinea pig에 투여한 결과 가장 미약한 열처리를 받은 70℃에서 15초 가열된 시료에 있어서도 생존하지 못했다. 고온단시간살균법에 의한 살균공정에서 결핵균의 사멸에 관한 실험결과(Harrington, 1965)에 의하면 저온장시간으로 살균된 717시료와 고온단시간으로 3,597 시료 중에서 결핵균 생균수 확인을 위한 guinea pig 실험결과, 감염율은 각각 1.1%와 0.03%로서 고온단시간 살균처리된 시료의 살균효율이 높은 것으로 확인되었다. Mycobacterium tuberculosis, M. bovis, M. avium과 M. fortitum 등을 균종별로 시험관 내에서 멸균된 탈지유에 오염시킨 후 62.8℃에서 30분, 71.7℃에서 15초 가열한 후 사멸효과를 확인한 결과 모두 사멸되었다.
이질(Shigellosis): 우유와 cheese가 원인이 된 이질의 발병이 보고된 적이 있는데, 우유 및 유제품에 보균자로부터 오염된 후 냉장하지 않은 상태에서 수 시간 방치된 제품을 섭취하여 발병된 것으로 확인되었다.
Shigella dyseteriae, Sh. flexerni, Shigella sonnei 등이 이질을 일으키는 병인균으로 가열에 의한 사멸조건을 조사한 결과 저온살균이 확실하게 수행된 경우, 이들 병원균은 사멸되는 것으로 확인되었다(Nevot et al., 1960).
Shigella spp.에 대한 열저항성에 관한 실험에서 우유에 2×107/ mL 수준으로 첨가한 후 62℃까지 예비 가열한 다음 70℃, 74℃, 80℃의 항온수조에서 15초간 가열하였다. 그 후 이 병원균의 생존성을 평가한 결과, 80℃ 이상 온도에서 15~20초간 열처리를 하면 Shigella sonnei가 사멸되었다고 보고하였다(Milyanovskii, 1970).
보트리눔증(Botulism): 미국에서 발생된 5건의 botulinum증 중에 2건이 우유와 연관되어 발병된 것으로, botulinum 증은 드물게는 Cottage cheese와 Neufachtel, Lieder kranz cheese 등에도 발생되었고, Yoghurt도 매개체로 작용되었을 것으로 인정되고 있다. Clostridium botulinum이 주요 병원균으로 포자(spore)를 형성하므로 열내성이 매우 높아 사멸시키기 위해서는 더 높은 수준의 열처리가 요구된다. Clostridium botulinum의 내열성은 혈청형 종류에 따라 다르게 나타나는데 A~G 6종의 혈청형 중에서 E형이 가장 내열성이 약하며 F형은 가장 내열성이 강하고, A형과 B형은 중간정도의 내열성을 나타낸다.
100℃에서 끓이는 정도의 열처리로 A형 혹은 B형의 포자를 불활성화 시키지는 못하는데, 통상적으로 저산성 식품에서 이 병원균 포자를 살멸하기 위하여 12D 처리를 추천하며 121℃에서 가열하는 것을 권장하며, Cl. botulinum의 D value는 100℃에서 100~ 330분이다.
Cl. botulinum 포자의 열저항성은 위에 제시한 바와 같이 큰 변화폭을 나타내고 있으며 열저항성에 영향을 주는 요인을 요약하면 다음과 같다.
Esty와 Myers(1922)에 의하면 배양 기간이 짧은 습도가 높은 상태의 Cl. botulinum 포자가 오래 배양된 상태의 포자보다 열내성이 높다. 또한 Sommer(1930)의 보고에 의하면 4~8일 배양 후의 Cl. botulinum 포자의 열저항성이 가장 높았다고 하였으며, 비포자 형성 세포 중에서도 열저항성의 변화를 보였다. 내열성 연쇄상구균에 속하는 미생물들은 성장곡선중 대수증식기 초기에 내열성이 가장 높았다고 보고하였다.
Williams 등(1937)은 멸균 또는 살균된 식육 중에서 생성된 포자의 내열성이 원료육에서 열처리 받지 않은 상태로 생성된 포자보다 약 3배 정도 내열성이 강한 것이 확인되었고 Sugiyama(1951)의 보고에 의하면 Cl. botulinum은 배지 중에 Fe2+ 및 Ca2+의 농도가 일정한 수준이하로 감소된 경우에는 포자의 열안정성이 감소되는 것을 확인하였다. 또한 장쇄지방산이 존재하는 경우 포자의 내열성은 증가한다고 보고하였다.
Amaha와 Odal(1957)은 망간과 칼슘이 Bacillus coagulans의 내열성을 증가시켰고 최대 내열성을 확보하기 위해서는 적정량의 망간 및 칼슘 등 2가 양이온이 적절한 수준으로 존재해야 한다고 밝혔다.
Sugiyama(1951)는 포자형성온도가 포자의 내열성에 영향을 미치며 고온에서 생성된 포자는 저온에서 생성된 포자보다 열내성이 높고, 또한 호기성포자의 경우 포자의 열내성이 최대생육온도와 연관되며 최대생육온도가 높은 포자의 내열성이 높다고 밝혔다.
자연적으로 생성된 포자는 인위적으로 유도시킨 포자보다 내열성이 높은 것으로 나타났다. Soberheim과 Mundel(1938)의 보고에 의하면 자연적으로 생성된 포자를 함유하고 있는 토양에서 인위적으로 유도된 포자의 경우보다 6~8배 장시간 열처리가 요구된다고 하여, 포자의 내열성은 배지의 물리적 상태에 따라서도 큰 차이를 보이는 것 같다. 이는 인위적으로 유도시킨 포자를 대상으로 실험한 결과가 실제 공장 환경에 적용시킬 때 차이가 나타날 수 있음을 시사하는 것이다.
가열 이후에 병원균이 배양될 회수용 배지의 조성이 가열살멸시간에 큰 영향을 미칠 수도 있다. Williams와 Reed(1942) 및 Sugiyama(1951)의 연구결과에 의하면 Cl. botulinum 포자를 열처리 한 후의 회수조건 및 배양온도에 따라 내열성에 영향을 미친다고 밝혔다. 즉 24℃, 27℃ 및 31℃에서 각각 배양된 Cl. botulinum포자가 37℃에서 배양된 것보다 회수율이 높았으며, 이러한 사실은 제품시험에 있어 중요한 의의가 있다.
가열 media의 pH는 내열성에 큰 영향을 미친다. Cl. botulinum에 있어서 pH 6.3~6.9의 범위에서 가장 높은 내열성을 보이는데, 대부분의 포자형성 미생물들은 중성 pH에서 내열성이 높다(Esty and Meyer, 1922).
수소이온농도의 증가는 통상 내열성을 감소시키는데, Lang(1935)은 Cl. botulinum 포자의 경우 pH 5.2~6.8 범위에서 pH와 내열성간의 상관관계는 발견할 수 없었다는 보고가 있으며, pH가 4.9일 경우에는 내열성의 급격한 감소가 확인되었다고 보고하였다. 또한 Xezones와 Hutchings(1965)는 식품 중에서 Cl. botulinum 포자의 pH에 의한 내열성 변화를 조사한 결과, 열처리 온도가 낮은 경우 pH의 영향이 내열성에 크게 미친다고 보고하였다.
Weiss(1921)는 Cl. botulinum 포자의 내열성이 염류를 3% 존재함에 따라 감소하였고, 염류농도를 증가함에 따라 열저항성은 급격히 감소한 것으로 발표되었다. 그러나 염류농도에 따른 효과가 이와는 상반되게 나타난 예도 있으며, Esty와 Meyer(1922)에 의하면 Cl. botulinum의 내열성은 염류가 1~2% 존재함에 따라 증가하였고 8% 이상으로 염류농도를 상승시킨 경우에는 내열성이 급격히 감소하였다. Cameron(1951)은 Cl. botulinum 포자의 내열성이 110℃ 이하의 온도에서는 염류에 의하여 열저항성 저하가 뚜렷이 나타났으나, 110~112℃ 온도범위에서는 뚜렷한 감소효과는 없었다고 하였다.
따라서 염류의 효과는 농도, 현탁용배지의 종류, 사용균주 등에 따라 가변적이며, 이들이 작용은 염류에 의한 pH의 변화 기타 용질의 해리정도의 변화에 기인된 것으로 판단된다.
당류, 그리세롤 기타 비독성물질은 미생물의 가열에 대한 사멸 효과에 관하여 연구한 결과, 이러한 물질이 충분한 농도로 존재할 경우 사멸 작용을 감소시키는 것으로 나타났다.
Sugiyama(1951)의 연구에 의하면 Cl. botulinum 포자의 열저항성은 설탕이 존재할 경우 증가하였으며 12.5% 정도의 낮은 농도로 설탕이 첨가되었을 때에도 약간의 사멸을 감소시켰다.
Gelatin이나 유청고형분 등 단백질물질도 미생물들에 있어서 열에 대한 보호작용이 있는 것으로 보고된 바 있으며(Anzulovic, 1932), Murray와 Headlee(1931)는 peptone과 같은 유기물의 양을 증가시키면 pH 7.0에서 C. tetani 포자의 가열사멸 시간을 연장하는 효과가 있음을 보고하였고, 혐기성 포자의 열저항성에 미치는 성장촉진 인자들의 효과에 관한 연구를 수행한 결과, 중성의 인산완충액에 100 ppm의 ascorbic acid를 첨가함에 따라 Clostridium spp.의 가열사멸시간을 24~36분 연장시켰고, Vitamin K를 첨가한 경우 열저항성 감소효과를 나타내었다.
여러 연구자의 연구결과를 종합하면, 지방이나 유지에 혼합되어 있는 병원균은 물에 혼합되어 있는 경우보다 가열 사멸이 어렵다고 보고한 바 있으며, Lang 등(1935)은 이러한 효과가 oil 매체의 낮은 열전도성에 기인한다고 주장했다.
Oil에 현탁된 Clostridium botulinum 포자는 이 미생물이 일반적으로 사멸되는 조건에서도 생존한다고 보고한 바 있으나, Nichols(1940)는 우유 또는 크림 중의 지방함량이 증가해도 Bacillus subtilis 포자의 내열성은 증가하지는 않았다고 하였다.
가열 media 중에 억제물질이 존재할 경우 가열저항성을 감소시킬 것으로 예측할 수 있다. Michener 등(1959)은 Cl. sporogens의 열저항성에 미치는 650종의 물질에 관하여 연구한 결과, ethyl oxide, diepoxy butane, 과산화수소, formaldehyde, dodecyl guanidine, hydrobromide, cetylmethylammonium bromide, 2․methyl-1,4-naphthoquinone, garlic oil 등은 열저항성을 감소키는 것으로 나타났다.
항생물질 중에 subtilin을 14 ppm으로 첨가하였을 때 Cl. botulinum과 PA 3679 포자의 열저항성의 변화는 대조구 대비 47%의 열저항성을 보였으며, Cl. botulinum 포자는 동일한 현탁액 조건하에서 대조구보다 63%의 열저항성를 나타내었다. O’ Brien(1956)은 pea puree에 subtilin과 nisin을 첨가한 경우 Cl. botulinum의 D value가 감소하였다.
항생물질은 포자의 열저항성에 영향을 미치지도 못하고 포자의 발아를 방지하지 도 않으며 발아 이후에 증식을 억제하는 물질로 작용하므로 억제할 수 있는 농도를 항상 유지해 주는 것이 매우 중요하다고 밝힌바 있다. 이러한 이유에 근거하여 항생 물질을 사용했더라도 Cl. botulinum 포자의 불활성화를 위한 허용된 최소 열처리수준 이하로 낮추면 안될 것이다.
E. coli 장염: Camiemberti cheese 및 Brie cheese가 Escherichia coli에 의한 장염(설사) 발병에 있어서 주요 원인균으로 보고된 바 있다.
특히 유아 중에서 이들 E. coli 장염 발생률이 높다. 사람이나 동물은 많은 수의 E. coli를 분변으로 배출하며, 이 미생물은 건강한 동물 젖이나 유방염에 걸린 젖소, 우유 제품 및 cheese에서도 종종 분리된다. 대장균군 중에서 열에 대한 내성이 강한 20균주를 분리한 다음 우유에 접종하고, 62.8℃에 가열하고 생존시간을 측정한 결과 1균주는 40분, 다른 1균주는 20분, 7균주는 10분간 생존하였다.
우유 중의 대장균군 가열사멸시간을 측정한 Olson 등(1952)의 연구결과를 보면, 분리된 139 균주의 대장균을 135°F에서 사멸에 도달하는 시간이 5분 이하부터 150분 이상까지 많은 편차를 나타내었다. Bacterium coli 형의 세균이 가장 내열성이 강한 것으로 확인되었고, 순수배양된 coliform의 가열 사멸시간은 오염된 균수가 많을수록 시간은 증가하였다. 병원성 E. coli 의 저온살균유에 존재 여부에 관한 실험에서 Papavassiliou(1957)는 영국에서 판매 되는 375시료의 저온살균유 중에서 병원성 E. coli가 분리되지 않았으므로 병원성 E. coli 는 저온장시간살균처리에 의해 사멸된 것으로 생각되었으며, 내열성 coliform은 약 3.7% 정도 존재하는 것으로 보고하였다. E. coli는 enterotoxin을 생산하는 균주들도 있지만, 독소를 생산하지 않는 균주도 있는데, 장점막에 E. coli 가 침범하거나 enterotoxin의 작용에 의하여 설사를 일으킨다.
E. coli가 생산하는 toxin의 가열에 대한 저항성을 측정한 결과, 60℃에서 5분간 가열한 이후 독성의 활성은 1/3로 감소하였고, 60 ℃에서 15분 가열에는 90%가 상실된 것으로 확인된 바 있다(Mundell, 1976).
Pseudomonas aeroginosa 장염: Pseudomonas aeroginosa는 열에 민감한 enterotoxin을 생산하며, 우유에 존재하여 장염을 일으키는 병원체로 어린이에게 빈번히 발병하는 것이 특징이다. 이 병원균은 Ps. fluoresence, Ps. putida와 함께 형광성 색소를 형성하는 특성이 있어 형광생성 Pseudomonas group으로 분류되며, 토양, 물, 퇴비 포유동물의 장내용물에 분포한다.
이들 병원성 미생물은 생육하면서 열안정성이 높은 phytotoxin과 열에 쉽게 파괴되는 Exotoxin A를 생성하며, 후자가 장상피에 괴저 및 출혈성 병변을 일으키는 것으로 확인되었다. 이 미생물은 55℃에 1시간 가열될 때 사멸된다(Parker, 1983).
리스테리아증(Listeriosis): Listeria가 식품을 통해서 인간에게 감염되는지에 관하여는 확인되지 않았으나, 원유가 하나의 원인이 될 수 있을 것으로 고려되고 있다.
Potel(1953)은 유방염에 걸린 젖소에서 착유된 우유를 음용한 임산부가 쌍태아를 조산하였고, 이 유산된 태아는 과립조직으로 이루어진 multiple tumor를 가지며 태아와 우유로부터 Listeria monocytogenes를 분리하는데 성공하였다.
Listeria증은 소에서도 발견된 바 있고 소에서 분리된 L. monocytogenes에 관하여 서술된 바 있다. 낙농 가공제품 중에서 L. monocytogenes의 내열성에 관한 연구를 수행한 Bradshaw 등(1987)의 보고에 의하면, L. monocytogenes Scott A균주를 105/ mL의 현탁액 상태에서 밀봉된 tube중에 52.2~79.4℃ 온도에서 여러가지 시간으로 가열하고 D value를 측정한 결과, 원유시료에서의 D value는 71.7℃에서 0.9초, 멸균유에서의 D value는 2.7초였다. Ice cream mix에서의 D value는 79.4℃에서 0.5초인 것으로 나타났다.
액상유제품에서 Z value는 5.8~7.1℃로 나타나 원유에 적정한 살균이 이루어지면 L. monocytogenes는 생존이 불가능한 것으로 확인된 바 있다.
54균주의 Listeria monocytogenes를 실험실 및 공장조건에서 내열성 실험결과 실험실 조건에서 이 균주의 우유현탁액을 모세관에 밀봉하여 65℃에서 35초, 75℃에서 10초, 85℃에서 20초 가열할 때 사멸되었고, 85℃에서 실시된 상업적 순간살균법으로 열처리한 결과 모든 균주가 사멸되었다(Dedie and Schultz, 1957).
Clostridium perfringens 장염: 어린이 설사의 원인으로 Cl. perfringens 장염이 기인된 경우가 있으며, C. perfringens는 우유에 쉽게 오염될 수 있고 그 포자는 저온살균처리에도 생존할 수 있을 뿐 아니라 저온살균 열처리로 인하여 포자를 gemination 시킬 수 있으며, 우유에는 미생물이 생육하기 좋은 많은 영양소를 함유하고 있기 때문에 문제를 일으킬 수 있다. 그러나 열처리된 우유를 냉장하면 포자와 영양형 세포의 증식을 방지할 수 있다.
C. perfringens의 내열성은 90℃에서 D value가 0.015분으로부터 8.71로 변화폭이 넓다. 또한 C. perfringens는 저온살균처리에 의해 살멸되지 않으므로 살균유에 있어서 미생물 최초 오염도를 측정하는 지표로 활용될 수 있다.
Bacillus cereus 장염: 우유와 아이스크림이 Bacillus cereus에 의한 장염의 발생원인으로 확인된 바 있다. 어린이에 있어 이 유형의 장염 발생 보고가 있으며, B. cereus의 오염원으로 목장에서 채취된 토양시료에 105~106/g의 B. cereus 가 존재하는 것으로 확인되었고(Labot et al., 1965), 목초 중의 오염도의 103~104/g 수준이며 세척된 우유통의 오염도는 102~106/cm2로 광범한 차이를 보였다.
Bacillus cereus의 내열성을 연구한 결과를 보면(Lonscu et al., 1966), 58개의 시료 중 42개의 시료 최초오염도가 10⁵/mL 수준이며, 저온살균 이후에도 80%는 생존하였다. Bacillus cereus 포자의 발아에 있어서 열처리의 영향을 조사한 결과, whey 첨가는 포자 발아를 촉진시켰고, HTST 저온살균도 포자의 발아를 촉진시키는 것으로 나타났다(Wilkinson et al., 1973).
Rajkowski(1987)는 B. cereus 17균주의 생화학적 및 생태학적 특성을 조사하였는데, 광물질이 제거된 물에서 100℃에서 D value는 0.6~27분이며 Z value는 7.4~14.5분이었다. 열에 의한 이 병원균의 사멸 효과를 조사한 결과 B. cereus의 오염 수준이 높을 때에는 UHT와 HTST 처리에 의하여도 생존가능성이 있다고 주장하였다.
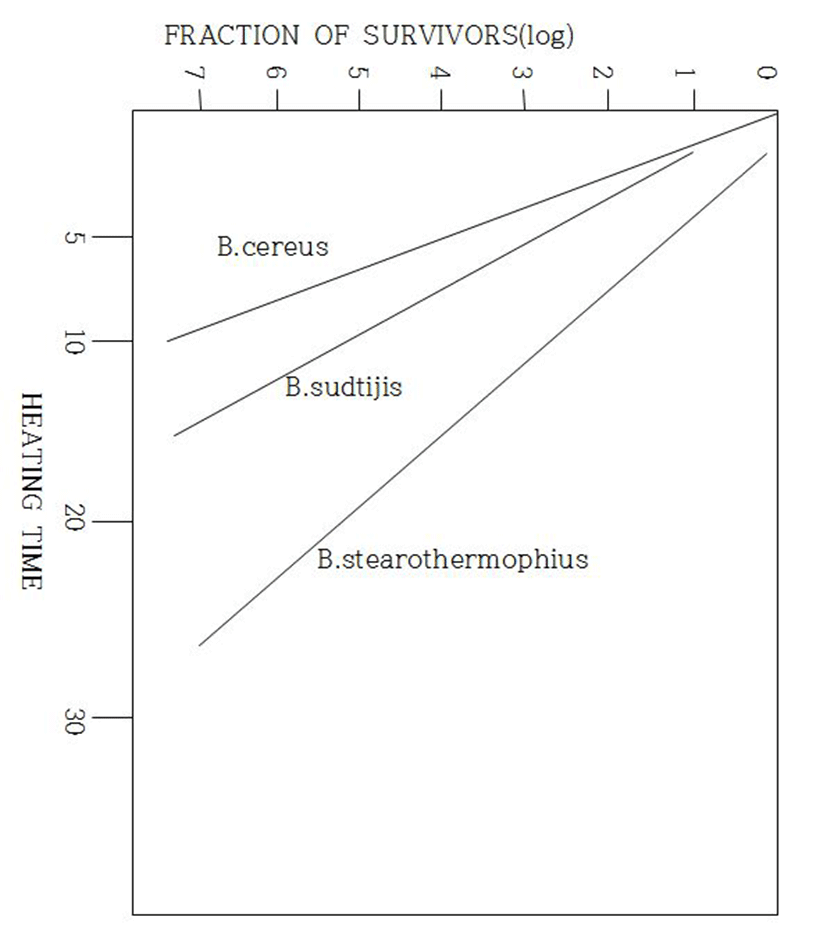
B. cereus를 포함한 호기성 포자형성 미생물의 물과 우유에서 사멸시간을 Table 4와 Table 5에 제시하였고, B. cereus와 다른 2균주를 Capillary 방법에 의하여 140℃에서 사멸성향을 조사한 결과를 Fig. 3에 제시하였다(Miller와 Kandler 1967).
Q fever: 원유에 존재하며, 병원체는 Coxiella burnetti 이다(Bryan, 1979). 원유가 이 질병을 매개한다는 근거는 보체(complement)를 결합시키는 항체 출현빈도가 생유를 전혀 마시지 않은 사람의 혈청에서 높게 나타난다는 사실에 근거한 것이다. 감염된 소로부터 착유된 생유를 마시는 것이나 동물, 먼지, 곤충에 접촉하는 것이 가능한 전염경로가 될 수 있다. Coxiella burnetti는 61.7℃에서 30분간 가열할 때에는 생존하므로 62.8℃에서 30분, 71.7℃에서 15초 정도 가열하면 사멸된다는 사실이 확인된 바 있다.
Enright 등(1957)의 조사결과, 우유시료 376개 중 14개 시료 중에 Coxiella burnetti 가 존재함이 확인되었고, 그 오염 수준은 최고 104 /mL이며, 오염된 생유를 143°F에서 30분 가열 시에는 사멸되지 않았으나 145°F로 30분 가열할 때 완전히 사멸하였으며, HTST 열처리에서는 161°F에서 15초간 가열해야 완전히 사멸한다고 보고하였다.
Aflatoxin 중독증: Aspergillus flavus 또는 Aspergillus parasiticus 가 식품 또는 사료에 존재하여 성장하는 과정에서 독소가 생산되는데, 이 독소를 섭취하는 경우 사람 또는 가축에 중독증상을 나타내게 된다.
미국의 경우 1978년에 우유에서 aflatoxin M1 오염사고가 보고된 예가 있으며, 1981년에도 이와 유사한 사고가 보고된 바 있다. Aflatoxin은 여러가지 유형이 우유 중에 나타날 수 있으나 aflatoxin B1이 수산화된 헝태인 aflatoxin M1형이다. 우유에 오염된 Aflatoxin M1(AFM1)은 시간경과에 따라 그 농도는 감소하여 0℃에서 4일이 경과하면 40%, 6일 경과 후에는 80%가 감소하였다(Mc-kinney, 1973). 그러나 5℃에서 1~3일간 저장하는 기간 중 11~25% 감소율을 나타내었다.
Purchase 등(1972)의 연구에 의하면 저온살균처리는 우유 중의 AFM1 함유농도를 저하시키며 열처리 온도가 높을수록 aflatoxin의 불활성화는 증가되며, 62℃에서 30분간 열처리할 때 불활성화된 양은 32%, 72℃ 및 80℃에서 45초간 HTST 처리할 때 불활성화된 독소함량은 각각 45%와 64%였다고 한다.
인위적으로 aflatoxin를 오염시킨 우유를 열처리한 경우에는 불활성화되지 않았다는 보고도 있어 주의가 필요하다. 한편, UHT 처리시에는 AFM1이 감소하는데 aflatoxin에 오염된 원유를 1 kg/cm2의 증기압(121℃)에서 20분간 처리할 때 80%의 aflatoxin 불활성화가 나타난 것으로 보고되었다(Purchase et al., 1972).
결론
우유 중에 오염되어 사람에 질병을 일으킬 수 있는 병원균들 중의 대부분은 비포자 형성균으로서 열처리에 의하여 사멸되므로 살균 공정은 유제품의 안전성 확보를 위해 긴요한 공정이다. UHT 처리를 할 때에는 포자형성균의 포자까지 사멸시키므로 안전성과 보존성 증진을 위하여 중요한 의미를 갖는다.
병원성 미생물의 내열성은 균의 특성, age, pH, 염류, 당류, 단백질, 지방, 억제물질 등의 존재에 따라 영향을 받는다. 우유 살균 과정에서 B. cereus와 Cl. perfringens는 완전히 파괴되지 않지만 우유의 저온 보존기간 중에 발아 및 증식하여 발병에 이르기에는 불가능할 것으로 판단된다.