
Introduction
Kombucha is a fermented beverage produced using black or green tea and a symbiotic culture of bacteria and yeasts (SCOBY). SCOBY is a complex consortium of lactic acid bacteria (LAB), yeasts and acetic acid bacteria (AAB) that generates organic acids, vitamins, polyphenols, and other bioactive compounds during fermentation [1]. The beverage has been associated with antioxidant, antibacterial, and anticancer effects, and the main organic acids (acetic, lactic, gluconic, and glucuronic acids) in kombucha contribute to sour flavor and functional properties of kombucha [2]. Fermentation of kombucha consists of two stages; aerobic first fermentation (20°C–30°C, 7–14-d, final pH 2.5–3.5) driven by the symbiotic metabolism of yeasts, AAB, and LAB within the SCOBY, and anaerobic secondary fermentation to enhance carbonation and flavor [3]. Although kombucha is traditionally produced from black or green tea, kombucha can be manufactured using diverse substrates such as herbs, fruit extracts, and plant materials, which significantly alter its bioactive profiles and functional properties [4].
Lotus root (Nelumbo nucifera) is a plant with low-caffeine (<1%). Lotus root has vitamin C, dietary fiber, starch, minerals, and various bioactive compounds including polyphenols, flavonoids, and polysaccharides [5]. The dominant phenolic compounds (gallocatechin, catechin, epicatechin, and gallic acid) in lotus have been proven to show antioxidant, anti-inflammatory, anti-obesity, anti-diabetic, hepatoprotective, and neuroprotective activities [6,7]. When lotus root is fermented, total phenolic and flavonoid contents are increased markedly resulting in enhanced antioxidant and anti-tyrosinase activity [8]. It seems that lotus root can serve as a promising low-caffeine functional ingredient for the development of novel fermented beverages such as kombucha [9].
Therefore, this study aimed to develop a low-caffeine functional lotus root kombucha (LRK) and to investigate the effects of the addition of Lactobacillus plantarum inoculation or yogurt starter culture to LRK on its physicochemical and sensory properties of LRK.
Materials and Methods
Roasted lotus root was provided by Daebon (Gyeong-san, Korea). SCOBY and starter broth were purchased from Slowoon (Korea), and white sugar (CJ Cheiljedang, Korea) were purchased commercially. Yoghurt stock culture (YC-380, Chr. Hansen, Denmark), consisting of S. thermophilus and L. delbrueckii subsp. bulgaricus, was provided by Pulmuone Danone (Korea).
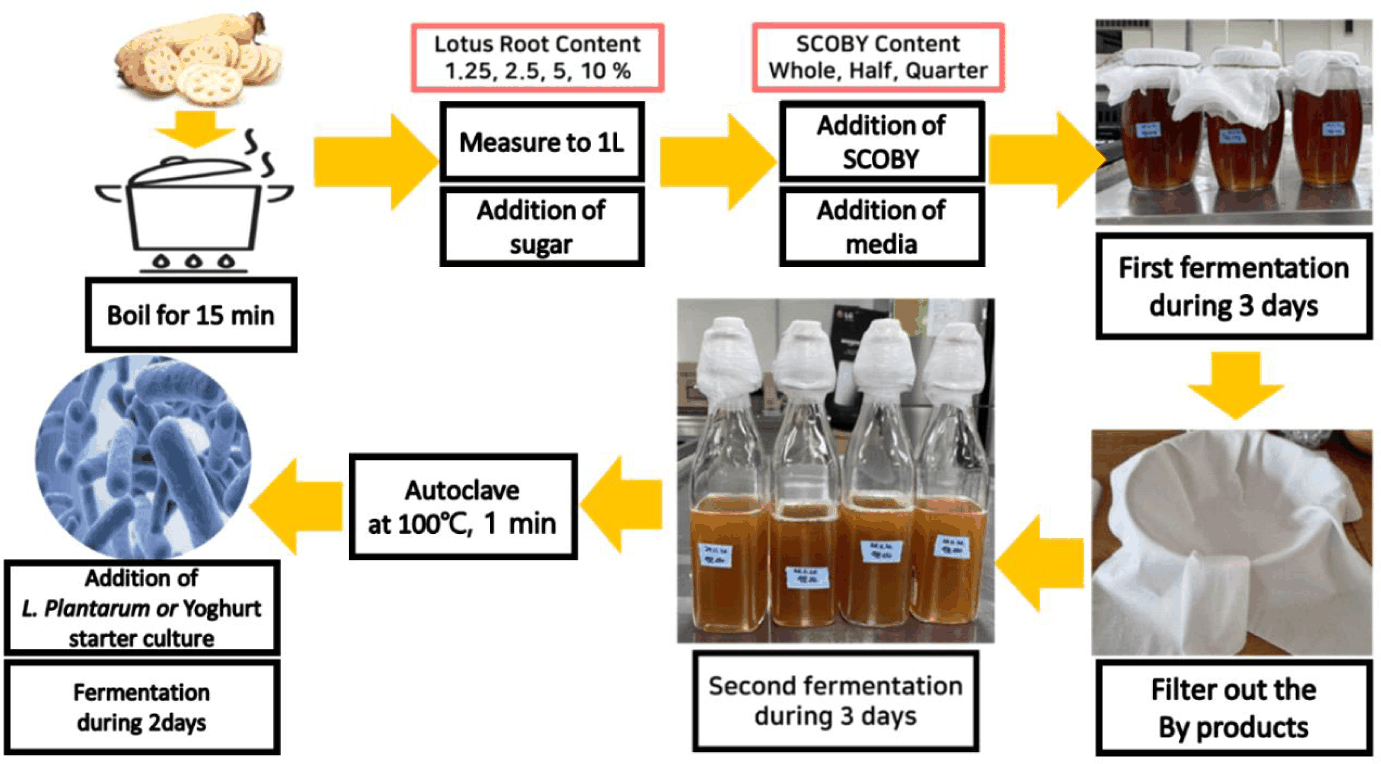
Lotus root was steeped in 1 L of water at 85°C for 15 min, and it was strained through cheesecloth. The steeped water was supplemented with sugar. After complete dissolution of the sugar, SCOBY (whole, half, and quarter) and 70 mL of broth was added. The first fermentation was conducted in a constant temperature and humidity chamber (DR-402, Daihan Scientific, Korea) at 25°C until pH was reached 3.8. After first fermentation, the product was strained through cheesecloth, and then it was sealed airtight, and it was subjected to secondary fermentation for 2 days at 25°C. After secondary fermentation, centrifugation was performed at 12,000 RCF for 5 min, and only the supernatant was recovered. After autoclave at 100°C for 1 min, Yogurt stock culture was inoculated at 0.2% (v/v) level or L. plantarum ATCC 14917 strain was inoculated with the concentrations of 0.2%, 0.5%, and 1.0% (v/v) into LRK, respectively (Fig. 1).
pH was measured using a pH meter (S220-K, Mettler Toledo, USA), and Brix was determined by using a digital refractometer (Digital Refractometer PAL-1, Atago, Japan). Moisture content was analyzed based on the Association of Official Analytical Chemists International (AOAC) method: samples were pre-dried at 80°C for 12 h and then dried in a drying oven (WiseVen Oven-N105, Daihan Scientific, Korea) at 105°C for 24 h. Moisture content (%) was calculated by using the following equation.
For L. plantarum inoculation, glycerol stocks stored at −80°C were subcultured twice in Man, Rogosa, and Sharpe (MRS) broth at 37°C for 24 h. The activated strain was recovered by centrifugation at 12,000 RCF for 1 min, then it was washed three times with sterile 0.9% (w/v) NaCl solution, and then it was washed once with distilled water. And then, activated strain was inoculated into kombucha at concentrations of 0.2%, 0.5%, and 1.0% (v/v). Storage stability was assessed every 7 days over a 28-day period at 4°C and 37°C, pH and Brix were monitored during 28-days storage.
Sensory evaluation was conducted with the approval of the Institutional Review Board (IRB) of Daegu University (approval no.: 1040621-202501-HR-015). A 7-point Hedonic scale (1: very dissatisfied, 4: average, 7: very satisfied) was used to evaluate color, odor, flavor, and overall acceptability by 10 trained panelists.
Data are reported as mean ± SD from triplicate independent experiments. For the evaluation of inter-group differences, one-way analysis of variance (ANOVA) was applied, with p < 0.05 set as the threshold for statistical significance. Computational analyses were carried out using the SAS software package, release 9.4 (SAS Institute, USA).
Results and Discussion
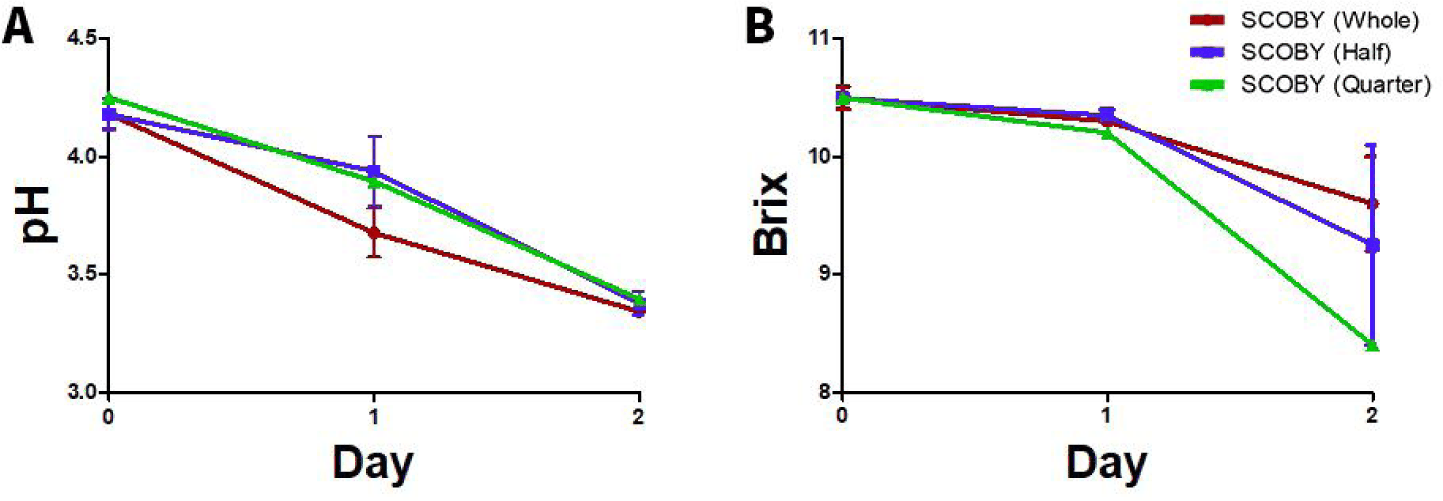
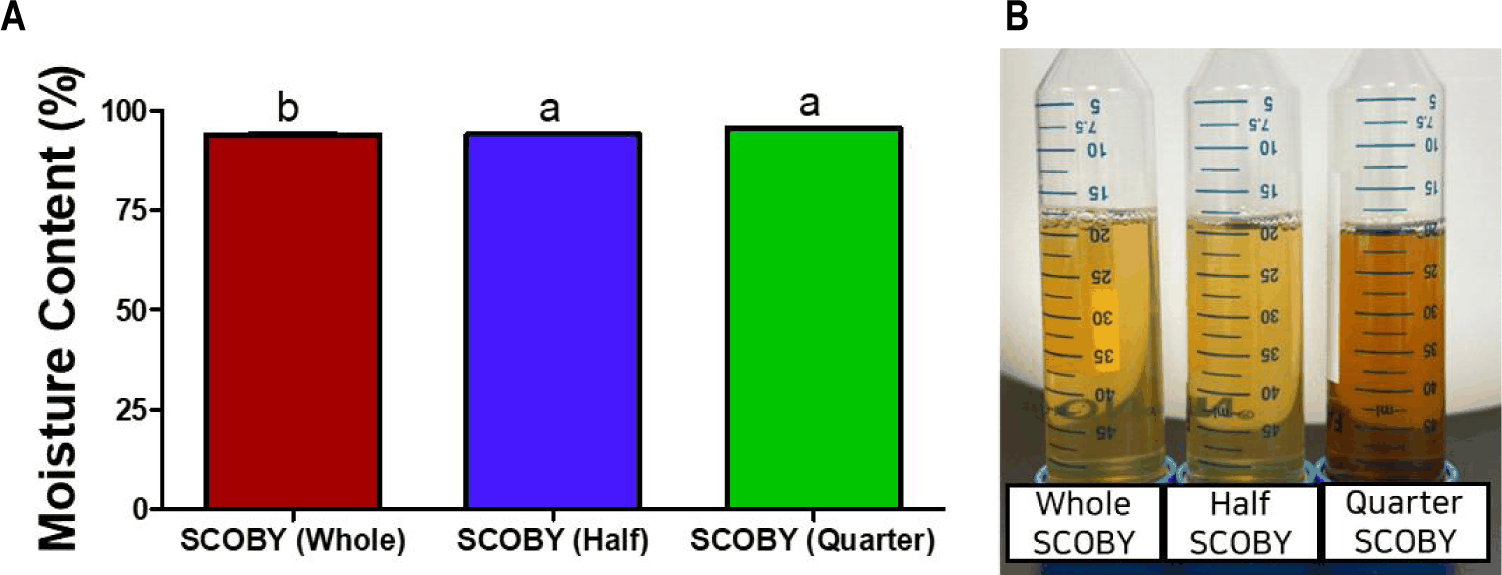
The changes in pH, Brix and moisture content during first fermentation of lotus root tea at three SCOBY inoculation levels (whole, half, and quarter) are shown in Figs 2 and 3. The moisture content (Fig. 3A) was significantly (p < 0.05) lowest when the whole SCOBY was used. This is likely because the whole SCOBY contains a higher amount of byproducts than other SCOBY content, which increases the total solid content. In all treatment groups, pH (Fig. 2A) was decreased significantly (p < 0.05) after fermentation. The whole SCOBY treatment showed the fastest acidification [10]. Brix (Fig. 2B) was also decreased after fermentation as SCOBY microorganisms convert sugars into organic acids, ethanol, and CO°C [11]. Similar trends have been reported in LRK, where higher SCOBY inoculation levels led to more rapid acidification and robust pellicle development[12]. Inoculum amount and microbial composition are primary determinants of acidification kinetics in kombucha, as increased inocula can consistently shorten fermentation duration by increasing the initial population density of metabolically active AAB and yeasts [13]. The proportional relationship between SCOBY mass and pellicle thickness further reflects the enhanced cellulose-synthesizing capacity of Komagataeibacter species [3]. It seems that SCOBY inoculation level is a decisive factor governing both acidification rate and sugar consumption during kombucha fermentation [14].
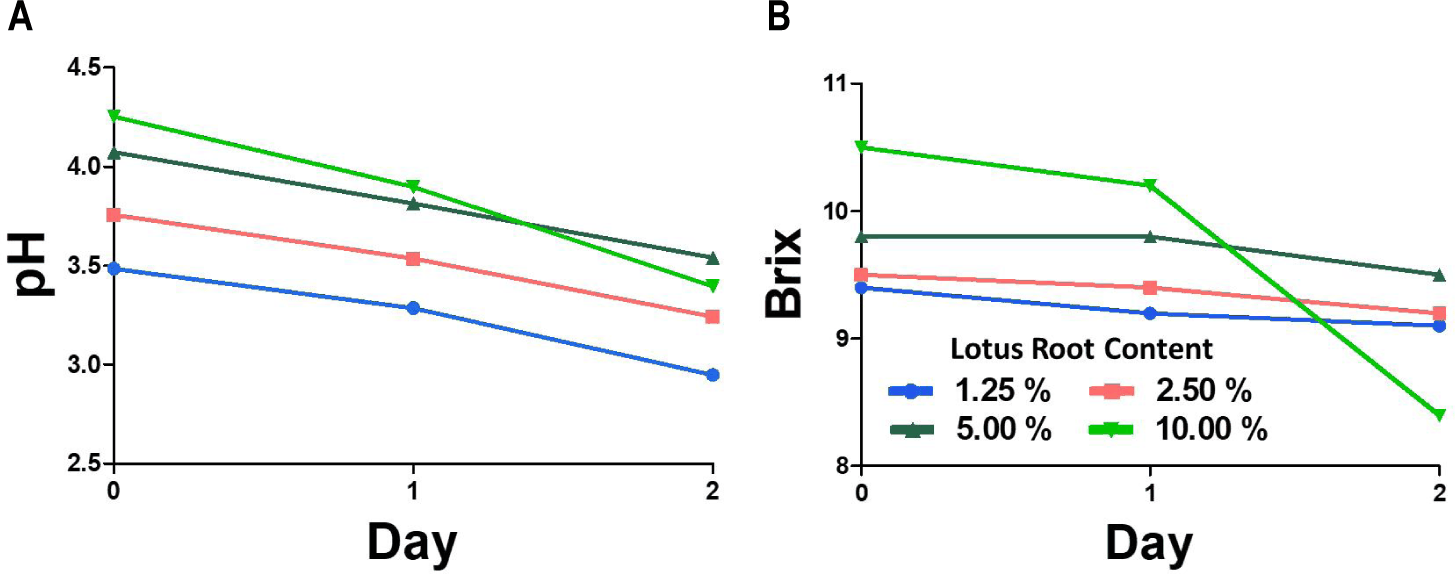
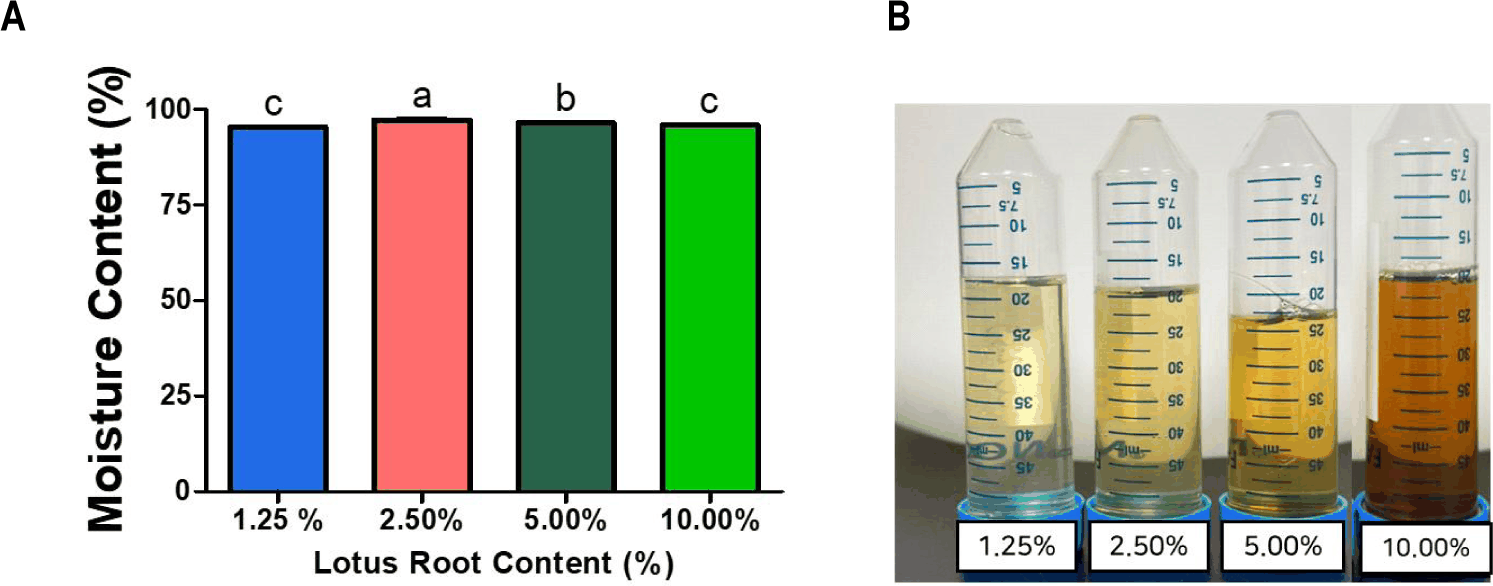
The changes in pH, Brix, and moisture content of LRK with four lotus root concentrations (1.25%, 2.50%, 5.00%, and 10.00% w/v) are shown in Figs 4 and 5. The pH (Fig. 4A) was decreased significantly (p < 0.05) at all concentrations after two days of fermentation [3]. Initial Brix (Fig. 4B) was increased when the used amount of lotus root was increased. After the fermentation, the Brix value was decreased variously among treatments, reflecting differences in the fermentable sugar pool available to SCOBY microorganisms [15]. Higher lotus root concentrations produced darker and more turbid fermentations, with brownish color most pronounced in the 10% treatment (Fig. 5A) [5]. The intensified browning at higher concentrations is likely due to the release and subsequent enzymatic and non-enzymatic oxidation of lotus root-derived phenolic compounds—such as catechin, epicatechin, and gallic acid—during fermentation. The color profile of LRK has been reported to be strongly associated with these intrinsic pigments and their oxidation products under fermentation conditions [12]. The physicochemical characteristics of plant-based substrates, including their polyphenol and soluble sugar content, have also been shown to meaningfully determine the metabolic environment of SCOBY microorganisms. Also, it ultimately accomplishes the bioactive profile of the resulting beverage [16]. These findings indicate that substrate concentration serves as an important processing variables governing both the physicochemical properties of kombucha and SCOBY metabolic activity during fermentation of kombucha [17].
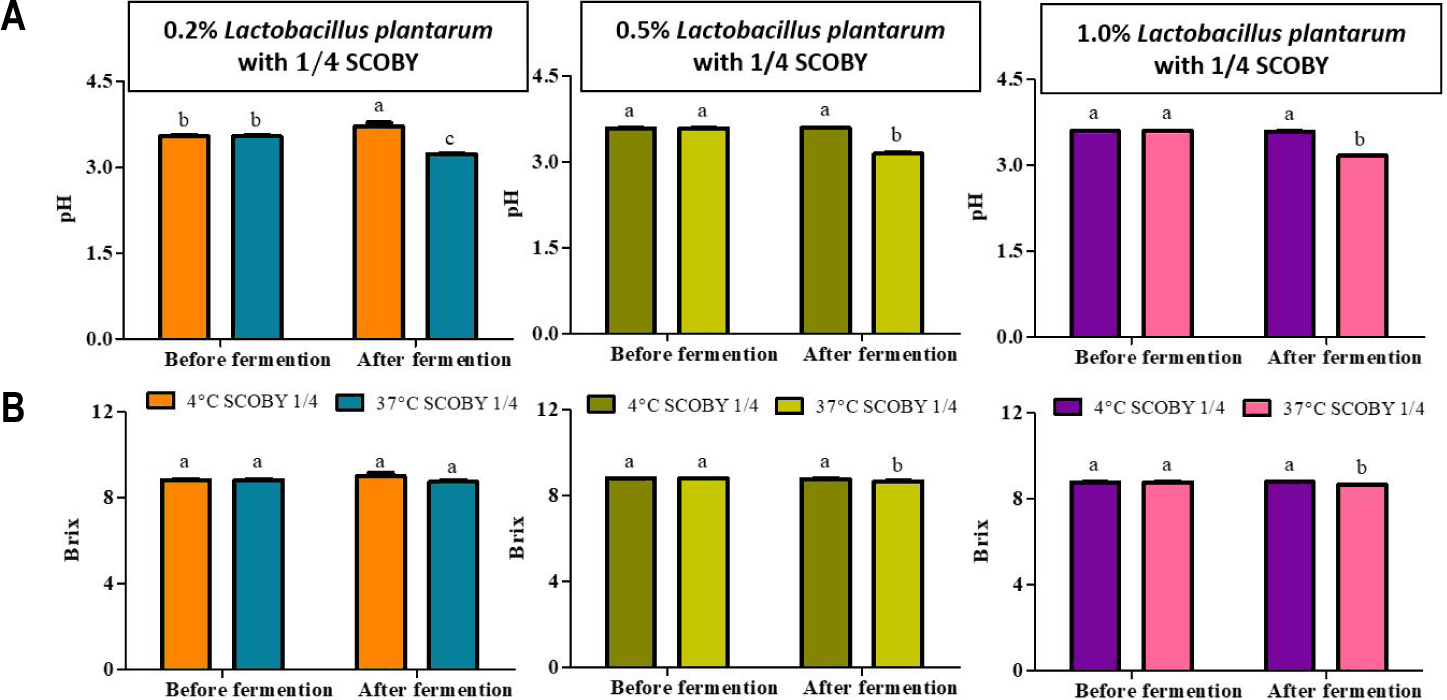
The changes in pH and Brix before and after secondary fermentation with L. plantarum (0.2%, 0.5%, and 1.0% v/v) at 4°C and 37°C are shown in Fig. 6. pH was decreased significantly (p < 0.05) at the three levels at 37°C [23]. This temperature-dependent pattern is consistent with the previous reports showing that temperature plays a key role in the growth rates and metabolic outputs of probiotic Lactobacillus strains in kombucha. It seems that lowering incubation temperatures markedly reducing acidification capacity by suppressing enzymatic activity and membrane-associated proton transport [24]. L. plantarum supplementation during kombucha fermentation has been shown to contribute to a significant accumulation of lactic acids and the present results also extend this finding to a lotus root-based matrix, suggesting that the low-caffeine lotus root does not interfere with L. plantarum metabolic function at permissive temperatures [20]. Secondary fermentation of LRK with L. plantarum at an inoculation level of 1.0 × 10°C CFU/mL has also been reported to increase lactic acid concentration and to enhance antimicrobial activity against Escherichia coli and Salmonella typhimurium, demonstrating that L. plantarum can exert meaningful metabolic activity in this specific lotus root environment [12].
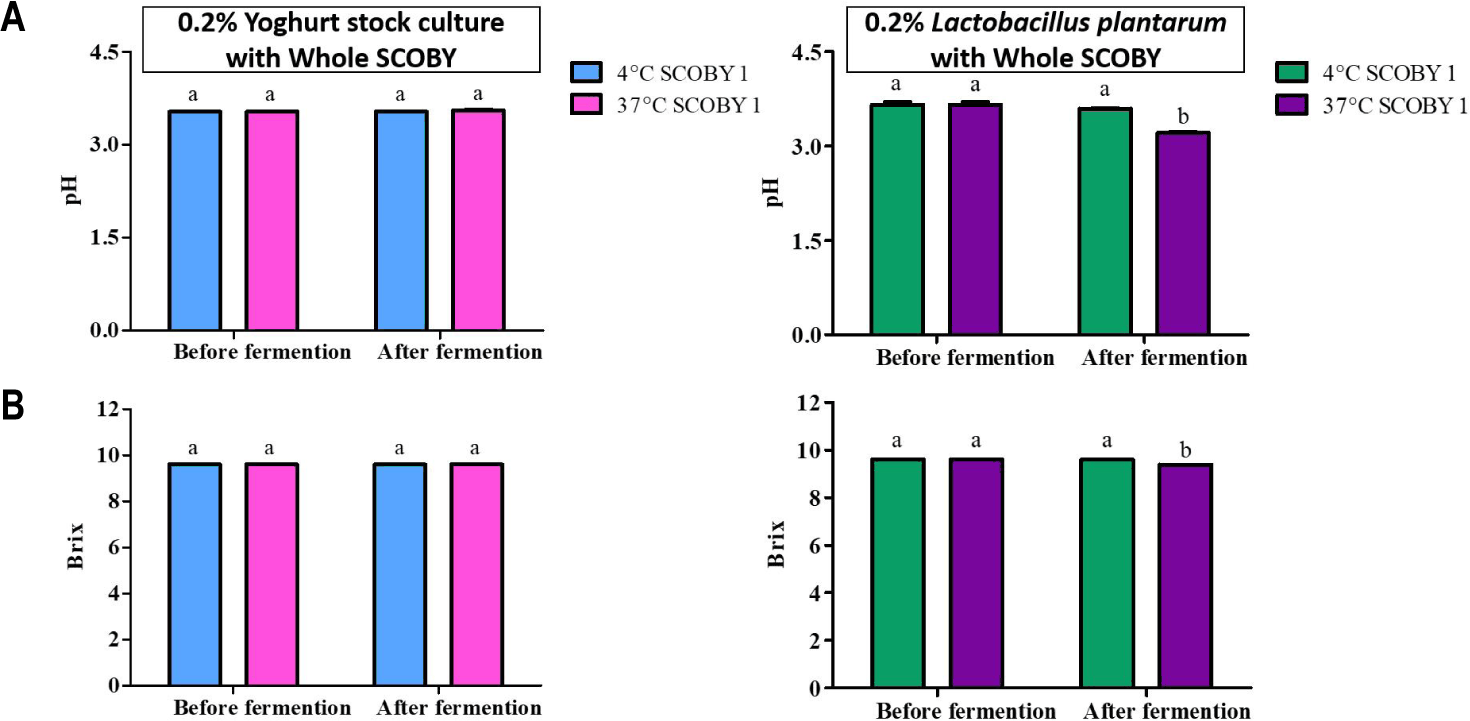
The changes in pH and Brix during secondary fermentation with yogurt stock culture (0.2% v/v) with different storage temperatures are shown in Fig. 7. No significant differences (p > 0.05) in pH or Brix were observed between two storage temperatures [22]. This result reflects the well-documented sensitivity of S. thermophilus and L. delbrueckii subsp. bulgaricus to low-pH, non-dairy environments; these strains are optimized for the growth in dairy matrices and typically lose viability and metabolic activity when exposed to the highly acidic conditions of kombucha [23]. It has been reported that the origin and physiological adaptations of LAB strains are critical determinants of their fermentation performance in kombucha, as strains are not acclimated to the acidic kombucha [25].
The changes in pH, Brix, and L. plantarum viable cell counts during 28-day storage at 4°C and 37°C w/ various inoculation level of L.Plantarum are shown in Fig. 8. At 4°C storage, pH and Brix were changed gradually throughout the storage period [21]. The viable cell counts at 4°C reached a maximum of approximately 1.0–2.0 × 10°C CFU/mL and was decreased gradually over 28 days. At 37°C, viable cell counts was dropped sharply after 1 week [24]. L. plantarum has been shown to remain viable in LRK at 4°C, supporting the practical suitability of refrigerated storage for probiotic LRK [12]. LAB-enriched kombucha samples have been reported to maintain viable counts (above 6–7 log CFU/mL) throughout fermentation at refrigerated temperature. It seems that temperature essential for preserving probiotic functionality in kombucha beverages [27]. The rapid decline of viable cell count at 37°C can be due to the combined effects of progressive organic acid accumulation, substrate depletion, and thermal stress on cell membrane integrity under elevated storage temperatures in kombucha [19]. These findings indicate that refrigerated storage at 4°C is the most suitable condition for maintaining L. plantarum viability in LRK [28]. The marked loss of viability at 37°C indicates that ambient-temperature is unsuitable for the survival in LRK. Therefore, refrigerated temperation and cold-chain distribution should be considered for the commercial production of LRK with probiotics , ensuring that adequate viable cell counts are maintained throughout the shelf life of product.
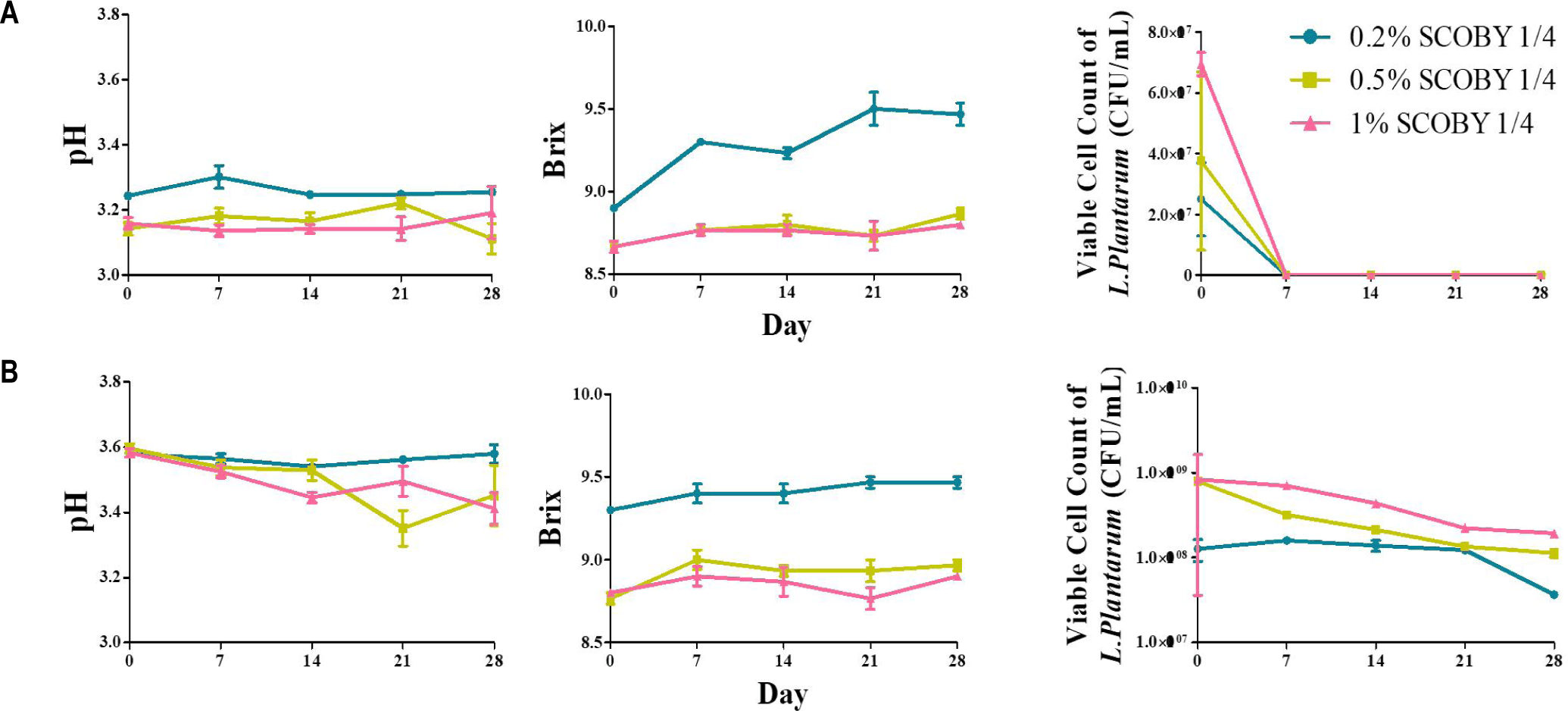
The pH, Brix, and viable cell counts of LRK supplemented with L. plantarum or yogurt stock culture during 28-day storage are shown in Fig. 9. In the L. plantarum group, pH was significantly (p < 0.05) lower at 37°C (3.18–3.25) than at 4°C (3.58–3.60), and Brix was increased gradually within the range of 9.4–10.0 in all groups over the storage period [26]. At 4°C, initial viable cell counts (~2.5 × 10°C CFU/mL) was decreased gradually to ~0.3 × 10°C CFU/mL by day 28; at 37°C, viable cell counts was below the detection limit after day 7 [20]. The continuing decline under refrigerated conditions is consistent with previous reports indicating that L. plantarum survival in kombucha fermentation broth is dependent on strain condition [20].
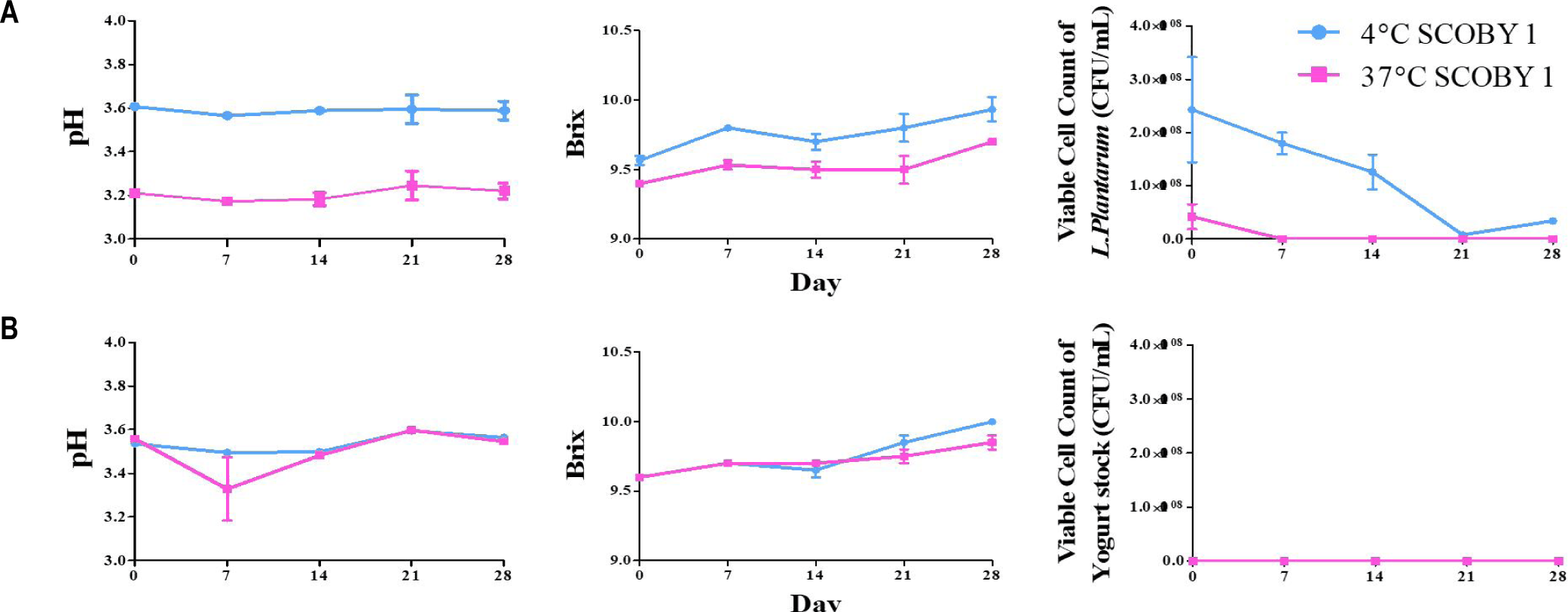
The rapid decreased of viable cell count at 37°C is attributed to the synergistic effects of acid stress and thermal pressure on cell membrane integrity in fermenting kombucha [19].
No viable cells were recovered from the yogurt stock culture group in both storage temperatures [22]. Compared with yogurt stock culture, L. plantarum was detected under refrigerated temperature (4°C), indicating survival in the acidic kombucha environment. It seems that yogurt stock culture bacteria, including S. thermophilus and L. delbrueckii subsp. bulgaricus, are adapted to the fermentation for the production of dairy products showing little survival when lactose is limited [23].
In contrast, L. plantarum, which is frequently isolated from fermented vegetables and other plant-based fermentations, exhibits tolerance to low pH and plant-derived phenolic compounds, allowing it to survive longer in kombucha during storage [25].
These results demonstrate that L. plantarum is a appropriate probiotic adjunct culture for LRK compared to yogurt stock culture.
The sensory evaluation results of LRK are shown in Fig. 10. Color was significantly (p < 0.05) highest in the whole SCOBY with 0.2% yogurt stock culture treatment. This result is consistent with the results that color in LRK is primarily governed by lotus root-derived pigments and the amount of SCOBY used [29]. Uncharacterized tea-derived pigment derivatives have been identified as the dominant determinants of kombucha color [13].
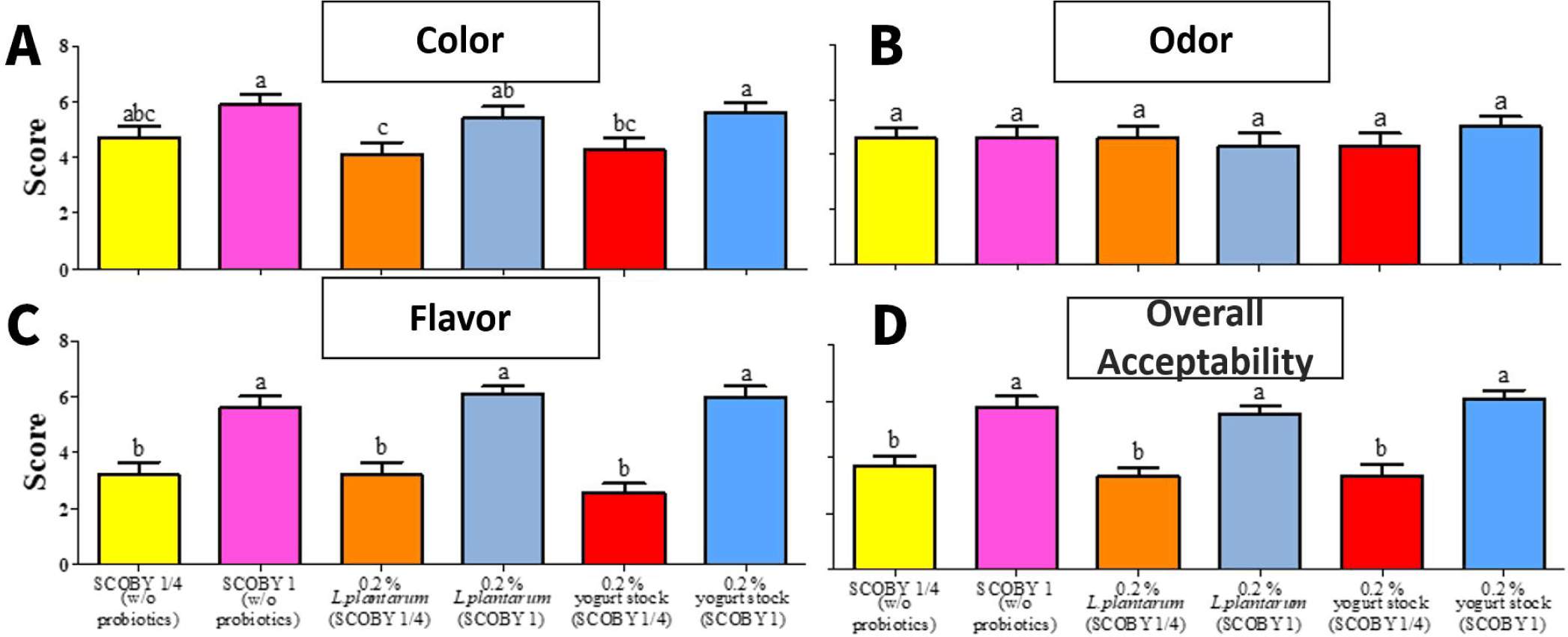
Flavor and overall preference were significantly (p < 0.05) higher in whole SCOBY samples regardless of the addition of probiotic [13]. The group without probiotics showed significantly (p < 0.05) lower flavor and overall acceptability scores compared to groups supplemented with L. plantarum or yogurt stock culture, indicating that LAB-derived fermentation metabolites contribute positively to the sensory quality of LRK [25]. Co-fermentation with L. plantarum has been demonstrated to improve the sensory quality of plant-based beverages through enhanced production of lactic acid and other organic acids, which positively correlate with sourness perception and overall acceptability ratings [30]. Supplementation of Lactobacillus during kombucha fermentation has also been shown to markedly elevate lactic acid content, resulting in a more complex and favorable flavor profile in the final beverage [20]. In addition, L. plantarum secondary fermentation in LRK has been reported to enhance antimicrobial and β-glucuronidase inhibitory activities alongside functional flavor-active metabolite production[12]. In the comparison between whole and quarter SCOBY preparations, the whole SCOBY group showed significantly (p < 0.05) higher overall acceptability, confirming that the amount of SCOBY is an important determinant of both fermentation kinetics and the accumulation of flavor-active compounds in LRK [13]. It is therefore concluded that both adequate SCOBY inoculation and probiotic supplementation are necessary conditions for producing LRK with optimal physicochemical and sensory characteristics. The sensory evaluation in this study was performed with a relatively small panel (n = 10), which may limit the generalizability of the findings. Although significant differences among the LRK samples were observed, further studies involving a larger and more diverse consumer panel are needed to confirm the sensory acceptability of the products.
Conclusion
It seems that lotus root is a promising low-caffeine substrate for kombucha production and that secondary fermentation with L. plantarum enhances the functional and sensory characteristics of LRK. In this study, fermentation conditions influenced product quality, where refrigerated temperature was essential for maintaining the viability of L. plantarum during storage. In addition, the use of whole SCOBY improved the sensory acceptability of LRK. Overall, the optimized fermentation process developed in this study provides a practical strategy for producing a probiotic LRK with improved consumer acceptability and potential commercial use. However, because the sensory evaluation was conducted with a limited panel of 10 participants, further intensive sensory evaluation is needed.
It seems that lotus root is an alternative to conventional tea for kombucha production. Also, L. plantarum can be effectively used as an adjunct culture to improve probiotic viability and sensory quality of the final product. Future studies should be needed validating the process at pilot scale on evaluating the effects of additional probiotic strains for the stability of LRK.