
서 론
구리(copper)는 체내에 필수적인 미량 원소이며, 다양한 생리학적 과정에 관여하는 여러 효소들의 보조인자로 작용한다. 구리는 항산화 방어 체계, 에너지 대사, 철 대사, 신경 전달, 콜라겐 합성, 색소 형성, 그리고 세포 발달 및 성장과 같은 필수 생리 기능에 관여하며, 세포 내 다양한 금속 단백질의 활성 조절에도 중요하다[1,2]. 구리는 주로 식이를 통해 섭취되며, 흡수는 소장의 상부에서 일어난다[3]. 체내 구리 항상성은 세포 내외의 구리 수송을 조절하는 다양한 운반체 및 조절 단백질에 의해 정교하게 조절된다. 대표적으로 CTR1(copper transporter 1)은 세포 내 구리 흡수에 관여하고, ATP7A(ATPase copper transporting alpha)와 ATP7B(ATPase copper transporting beta)는 구리의 세포 내 분포 및 배출을 담당하며, 과잉 구리는 간을 통해 담즙으로 배출된다[4]. 반추동물에서는 구리 항상성이 쉽게 교란될 수 있으며, 특히, 구리는 몰디브덴, 황과의 길항작용을 통해 흡수가 조절된다. 이들 미네랄은 thiomolybdate라는 복합체를 형성하여 구리의 흡수를 방해하고, 체내 구리의 조효소 기능을 억제한다[5]. 그 결과 생리적․병리적 이상이 나타나고, 다양한 여러 연구에 따르면 구리 항상성 붕괴는 반추동물에서 체중 감소, 발육 부진, 간 기능 이상, 모의 각질화 그리고 면역 기능 저하 등을 유발한다[6–8]. 이러한 이유로 구리의 적절한 섭취와 항상성 유지는 반추동물의 정상적인 성장과 면역 기능 유지, 생산성 향상을 위해 매우 중요하다.
반추동물 중에서도 특히 젖소에서 만성 구리 중독 사례가 여러 국가에서 보고되고 있으며, 그 발병 빈도는 점차 증가하는 추세다[9,10]. 영국, 네덜란드, 미국 등에서 다수의 젖소가 간 조직 내 구리 농도가 생리적 범위를 초과한 상태로 도태되고 있으며, 이는 만성 구리 독성에 노출되어 있음을 시사한다[9,11,12]. 특히 홀스타인(Holstein) 및 저지(Jersey) 품종에서 이러한 현상이 두드러지게 나타났으며, 이는 목초만으로는 충분한 구리 공급이 어렵기 때문에 외부 사료를 통한 과도한 구리 보충이 주요 원인으로 지목된다[12,13]. 이와 같은 문제는 반추동물 구리 항상성의 중요성에 대한 인식 부족과 제한적인 이해에서 기인한 것으로 사료된다.
젖소는 임신과 분만을 거치며 유선의 구조적․기능적 발달을 통해 우유를 생산하게 된다. 유선 발달 과정에서는 유선 상피세포의 증식과 분화가 활발히 이루어지며, 적절한 구리 수준 유지는 유선 상피세포의 발달에 필수적이다[14,15]. 하지만, 과다한 구리 수준은 활성 산소종의 생성을 유도하고, 산화 스트레스를 증가시켜 우유 생산성을 저하시킬 뿐만 아니라 유방암 유발 가능성까지 높일 수 있다[16,17]. 이러한 구리 과잉은 결과적으로 우유 생산량 감소, 유질 저하, 생식 효율 악화 등 생산성 전반에 부정적인 영향을 초래하므로, 젖소의 생체 내 구리 수준을 적절히 유지하는 것이 중요하다. 따라서, 젖소의 상태를 주기적으로 모니터링하고, 혈중 구리 농도 측정 및 조직 생검을 통해 체내 적정 구리 수준을 유지하는 관리가 필요하다.
이에 본 총설은 반추동물의 구리 항상성 조절 기전을 제시하고, 이를 바탕으로 젖소의 비유 생리에서의 중요성을 고찰하며, 나아가 낙농 산업 전반에 미치는 영향을 논의하고자 한다.
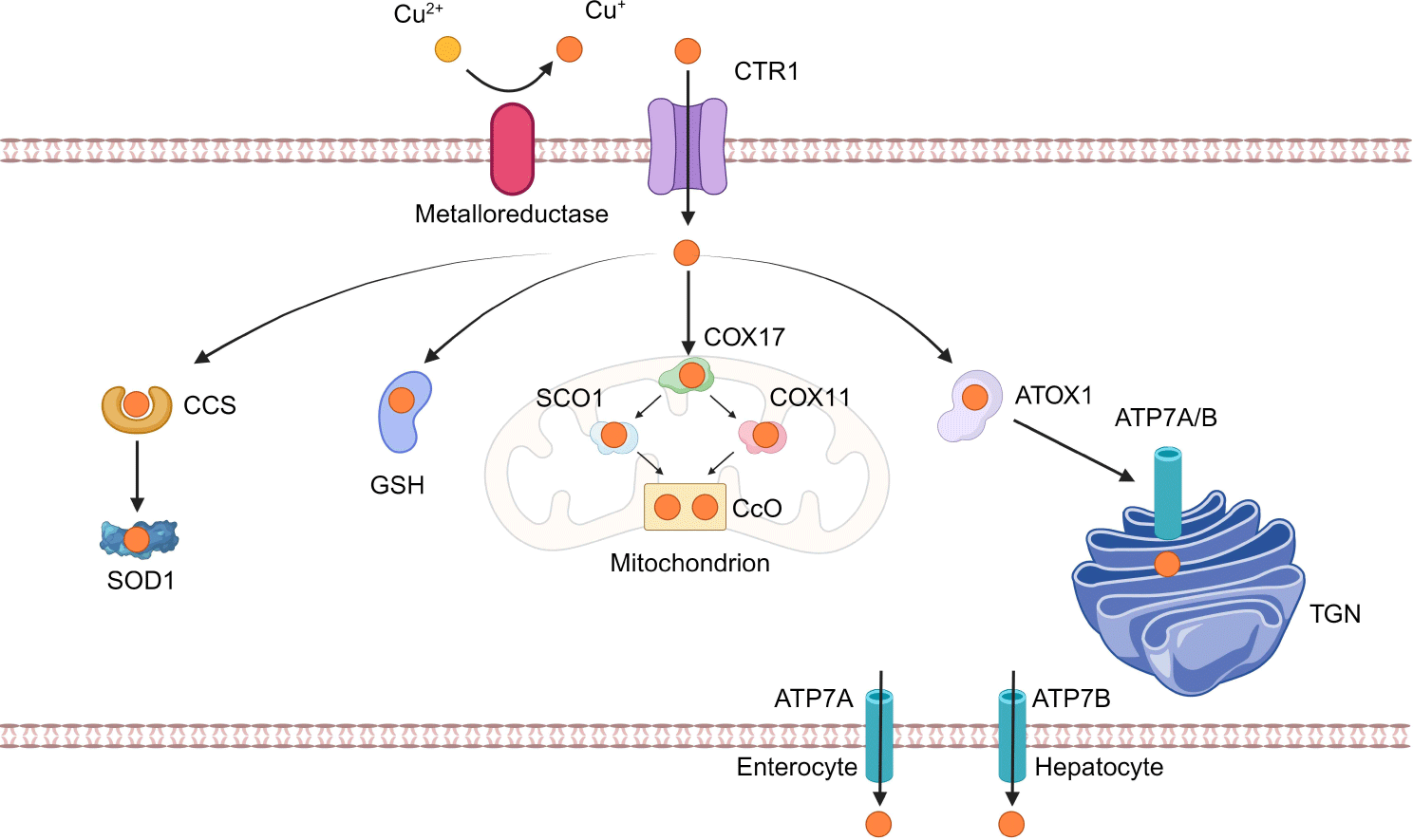
구리는 반추동물을 비롯한 모든 생물종의 생명 유지에 필수적인 미량 원소로, 다양한 생리적 과정에 관여하는 효소들의 조효소로 작용한다. 구리는 식이를 통해 공급되며, 장 상피세포에서 흡수된 뒤 ATP7A에 의해 혈류로 방출된다[1]. 방출된 구리는 albumin과 α2-macroglobulin과 같은 수용성 단백질에 결합하여 간으로 운반된다[18]. 이후 세포 외 구리는 CYBRD1(cytochrome b reductase 1)과 같은 metalloreductase에 의해 일가 구리 이온(Cu+)으로 환원되고, 원형질막에 존재하는 고친화성 구리 수송체 CTR1를 통해 세포 내로 유입된다. 세포질로 들어온 구리는 구리 샤페론 단백질과 결합하거나, 금속결합 단백질인 metallothionein에 저장된다. 간세포 내 구리가 과잉일 경우 ATP7B가 구리의 배출을 촉매하여 담즙을 통해 배설되도록 한다[19,20].
세포 내로 유입된 구리는 다양한 구리 의존 단백질의 조절을 통해 항상성이 유지된다. 예를 들어, ATOX1(antioxidant 1 copper chaperone)과 CCS(copper chaperone for superoxide dismutase), COX17(cytochrome c oxidase copper chaperone)과 같은 구리 샤페론, 또는 glutathione과 같은 비단백질 리간드에 결합하여 세포 내에서 운반된다. ATOX1은 구리를 ATP7A와 ATP7B로 전달하여 lysyl oxidase, tyrosinase, ceruloplasmin 등 구리효소의 합성을 촉진한다. CCS는 구리를 SOD1(superoxide dismutase 1)에 전달하여, 구리의 산화환원 반응에서 생성되는 활성산소종을 제거하는 항산화 작용에 기여한다. COX17은 미토콘드리아 기질에서 전자전달계의 복합체 IV인 CcO(Cytochrome c Oxidase)에 구리를 전달함으로써 세포 에너지 생성에 중요한 역할을 한다(Fig. 1)[6,19,20].
구리는 반추위에서 황과 몰리브덴의 결합으로 생성된 thiomolybdate와 복합체를 형성한다. Thiomolybdate는 체내로 흡수되어 구리와 결합함으로써 구리의 가용성을 크게 저하시킨다[21,22]. 소화관 내에서 thiomolybdate는 이용 가능한 구리와 결합하여 불용성 침전물을 형성하지만, 충분한 구리가 존재하지 않으면, 구리와 함께 침전이 되지 않은 자유 thiomolybdate가 전신 순환계로 흡수되어 구리 샤페론과 효소 등에 결합된 구리를 표적으로 하여 이들의 활성을 감소시킨다[5,6,23]. CcO와 SOD1과 같은 구리 의존 효소들은 에너지 생성 및 항산화 작용에 필수적이나, thiomolybdate가 해당 효소의 구리에 결합하면 이들의 활성이 저하된다[24,25]. 이는 구리를 완전히 제거하지는 않지만, 효소의 기능을 억제하는 것으로 알려져 있다[21,26,27]. 이러한 영향은 가역적이며, thiomolybdate 형성 조절이 구리 의존 효소 활성을 유지하는 데 필수적임을 시사한다.
반추동물은 담즙을 통한 구리 배설 능력이 매우 제한적이며, thiomolybdate에 의해 구리 흡수가 저해되어 구리 항상성 조절에 어려움을 겪는다. 구리 결핍을 예방하기 위해 사료 내 구리 보충량을 증가시키면 건물질 내 총 구리 농도는 높아지지만, 동시에 몰디브덴, 황, 철과 같은 구리 길항제의 농도도 함께 상승하여 오히려 2차 구리 결핍을 초래할 수 있다[20,28]. 예를 들어, 철과 구리의 결합은 thiomolybdate-구리 복합체 형성에 필요한 구리의 양을 감소시키며, 그 결과 자유 thiomolybdate가 체내로 더 많이 흡수되어 구리 결합 단백질의 활성을 억제한다[5,29]. 이를 방지하기 위해 구리 농도를 더욱 높이고 구리 길항제를 줄일 경우, 만성 구리 독성이 발생할 위험이 있다[20]. 실제로 전 세계적으로 특히 낙농우에서 구리 독성 사례가 증가하고 있으며, 홀스타인과 저지 품종이 가장 큰 영향을 받는다[12,13]. 이러한 구리 항상성 붕괴는 반추동물 구리 대사에 대한 이해 부족과 구리 항상성의 중요성에 대한 인식 부족에서 기인한 것으로 보인다.
구리 항상성의 붕괴는 반추동물 건강에 심각한 생리․병리적 영향을 미친다. 구리 불균형은 다양한 효소의 활성을 저해하고, 종을 불문하고 여러 유전 질환을 유발한다[6,30]. 인간에서는 Menkes 병과 Wilson 병이 대표적인 구리 관련 질환으로, 발달 장애, 간 기능 부전, 모발 변화 등의 증상을 보인다[31]. 반추동물에서도 Wilson 병과 유사하게, 간 내 구리 축적이 세포 핵 구조의 붕괴를 유도하여 세포 사멸을 초래하고, 막 지질 과산화로 인한 세포 괴사가 보고된 바 있다[32,33]. 또한, 구리 항상성 이상은 세포 발달과 성장에 큰 영향을 미친다. 여러 연구에서 구리는 유선 상피세포 발달과 호르몬 분비에 필수적이며, 과잉 공급 시 유방암 발생에 기여할 수 있음이 보고되었다[14,16,17]. 반면, 적정 수준의 구리 공급은 유방염 원인 미생물의 성장을 억제하여 유방염 발생률을 낮출 수 있으며[34,35], 젖소에서 구리 항상성 유지 여부는 단순한 대사 균형을 넘어, 유방 조직의 구조적 완전성과 분비 기능, 나아가 지속적인 젖 생산과 직결된다. 이에 따라 젖소 사양 관리에서 구리의 적정 농도를 정밀하게 조절하는 것은 생산성과 경제성을 동시에 확보하기 위한 핵심 전략으로 평가된다.
젖소의 유선 발달은 자궁 내 발달, 출생기, 이유기, 발육기, 임신기, 그리고 유즙분비기에 걸쳐 단계적으로 진행되는 복합적인 생리학적 과정으로, 호르몬, 영양 상태, 유전적 요인 및 환경적 조건이 정밀하게 상호작용하며 조절된다. 유선 발달은 임신 약 한 달째 자궁 내에서 시작되며, 작은 관과 유선 지방조직이 형성되고 유두가 완성된다. 출생 시에는 이미 기본적인 관상 구조가 갖추어져 있으며 기능적 활성은 없는 상태이다. 출생기와 이유기 동안에는 관 구조의 확장과 증식이 활발히 진행되고, 성장호르몬의 작용으로 발달이 가속화된다[36,37]. 사춘기에는 에스트로겐과 프로게스테론 분비에 의해 관 구조가 지방조직 속으로 침투․확장하고, 유선 상피세포 발달이 촉진되면서 유선 성장이 활발해진다. 임신기는 유선 발달의 결정적인 시기로, 에스트로겐, 프로게스테론, 프로락틴 등 호르몬의 작용에 따라 소엽-포상 구조 발달이 본격적으로 개시된다. 이 시기에는 단백질, 지방, 유당 합성능력이 급격히 증가하고, 포상세포가 유즙 분비를 위한 생화학적 및 구조적 분화를 진행한다[38,39]. 분만 후에는 프로게스테론 농도가 감소하는 반면 프로락틴, 코르티솔, 성장호르몬이 우세해지면서 유선 상피세포의 급격한 증식과 함께 본격적인 젖 성분 분비가 이루어진다[40,41]. 수유기에는 지속적인 착유 자극이 옥시토신 분비를 유도하여 젖 방출을 촉진하며, 장기간 수유가 우유 방출을 억제할 수 있고 이후 건유기를 거치면서 유선은 재생과 재분화를 통해 회복되어 다음 발달 주기를 준비한다[41,42].
유선 발달과 우유의 질은 호르몬뿐만 아니라 영양 상태에도 크게 의존한다. 에너지와 단백질 급여 수준뿐 아니라 비타민과 미네랄은 세포 구조 유지와 효소 활성에 필수적이다. 세포 수준 연구에 따르면, 휴지기–수유기–포유기로 진행됨에 따라 CTR1 발현이 증가하고, 이에 따라 구리의 축적도 증가하는 것으로 나타났다[14]. 또한, 적정 수준의 구리 첨가는 세포 주기 관련 유전자 활성과 항산화 작용을 통해 유선 상피세포의 증식을 조절하며[15], 이는 정상적인 우유 생산을 위해 구리 항상성이 유지되어야 함을 시사한다. 그러나, 과도한 구리는 유선 상피세포 내 산화 스트레스를 유도하여 세포 사멸을 촉진하고, 단백질 및 지질의 생합성을 억제함으로써 유선 세포의 증식과 기능을 저하시켜 우유 생산에 부정적 영향을 미친다[16]. 실제로 구리는 유방암 발생과도 밀접한 관련이 있으며, 연구에 따르면 구리는 유방암 세포에서 미토콘드리아 산화적 인산화 기반의 에너지 공급을 통해 암세포 성장을 촉진시킨다. 그러나, 미토콘드리아 내 구리의 결핍은 에너지 생성을 저하하여 세포사멸을 유도하고, 궁극적으로 종양 성장을 억제하는 것으로 나타났다. 또한 다른 연구에서는 유방암 환자의 조직과 혈청에서 구리 농도가 유의미하게 상승해 있음이 보고되었고, 이는 구리가 유방암 발달과 진행에 중요한 역할을 할 수 있음을 시사한다[17,43]. 이러한 현상은 인간뿐 아니라 젖소에서도 유선 조직의 섬유화, 종양 형성과 같은 병리적 변화로 나타날 수 있다. 이러한 이유로 수유기 젖소의 구리 권장 섭취량은 과거보다 크게 낮추어 설정되었으나, 산업화로 인한 토양 및 수질 내 구리 오염은 젖소에서 만성 구리 중독을 유발하고, 실제 섭취량이 권장 기준을 초과할 위험을 높인다[20,44]. 따라서 환경적․유전적 요인에 따른 구리 축적은 유선 세포 손상을 초래하고 유선 발달을 방해하며, 이는 우유 생산성과 품질에 잠재적인 위협이 될 수 있다. 결과적으로 젖소의 구리 항상성 유지와 이를 통한 유선 기능 조절의 중요성이 강조된다.
젖소의 건강을 유지하기 위해 구리 항상성 조절의 중요성이 강조되고 있으며, 이를 기반으로 한 체계적인 건강 관리 전략이 필요하다. 구리의 흡수와 배출은 일반적으로 체내에서 엄격히 조절되지만, 반추동물은 구리를 저장하는 능력 및 담즙을 통한 구리 배설 능력이 제한적이기 때문에 외부적 요인을 통한 흡수 조절이 필수적이다[19]. 이를 위해 사료 내 구리 함량과 구리 길항제의 수준을 환경 조건과 사양 관리 상황에 맞추어 조절해야 한다. 특히 구리는 사료뿐만 아니라 토양, 수질, 사육 환경에서도 축적․노출될 수 있으므로, 환경 내 구리 농도가 높은 경우 사료 내 구리 급여량을 낮추는 전략이 필요하다[20]. 또한, 젖소의 생리적 단계와 계절적 요인에 따라 구리 요구량이 달라지므로, 품종별․개체별 특성을 고려한 정밀한 영양 관리가 요구된다. 실제로, 저지 종은 홀스타인 종보다 구리에 덜 민감하다[12,13]. 이는 품종 간 차이를 고려한 구리 관리 체계의 필요성을 시사한다.
사료를 통한 구리 항상성 조절이 관리의 첫걸음이라면, 정기적인 영양 상태 모니터링은 그 다음 단계라 할 수 있다. 구리 결핍 혹은 중독은 체중 감소, 행동 이상, 발달 지연 등 외부적으로 관찰 가능한 증상으로 추정할 수 있으나, 면역력 저하, 간․심장․신경계와 같은 조직 특이적 손상, 암 발생 위험 증가 등은 단순 관찰로는 파악하기 어렵다. 따라서 혈중 구리 농도, 혈청 ceruloplasmin 수준, 구리 의존 효소 활성도와 같은 생체 지표를 이용한 정밀 진단이 필요하다.
구리 항상성 유지의 중요성이 젖소를 포함한 반추동물의 생리․병리학적 관점에서 강조됨에 따라, 구리 대사의 이해와 개체 및 품종별 민감도를 반영한 정밀 관리 전략이 필요하다. 이를 위해서는 사료 내 구리와 구리 길항제 함량뿐만 아니라, 사육 환경과 개체의 건강 상태를 고려한 종합적이고 맞춤형 관리 지침이 요구된다. 더 나아가, 반추동물에서 구리 대사의 분자적 조절 기전에 대한 심층 연구는 향후 구리 영양 관리의 효율성을 높이는 중요한 토대를 제공할 것으로 기대된다.
낙농 산업은 매년 꾸준히 성장하고 있으며, 전 세계 우유 생산의 연평균 증가율이 이를 뒷받침한다. 그러나 유제품 소비가 지속적으로 증가할 것으로 전망되는 반면, 인플레이션에 따른 소비 둔화, 환경 문제, 그리고 가축의 건강 상태는 여전히 중요한 도전 과제로 작용한다[45,46]. 본 총설에서는 젖소 건강 관리에서 구리 항상성 조절의 중요성을 고찰하였으며, 이는 낙농 산업 전반의 생산성과 경제성에 직접적으로 연결될 수 있음을 강조한다.
구리의 적정 공급은 유선 상피세포의 구조적 안정성과 기능을 최적화하여 우유 생산량과 품질 향상에 기여한다[15,35]. 특히 산화 스트레스가 최소화됨으로써 장기적인 착유 효율이 개선되고, 수유 기간 동안 안정적인 생산이 가능해진다[16,47]. 이는 농가 차원에서 생산 변동성을 줄이고, 제품 품질을 일정하게 유지하는 데 큰 도움이 된다. 더 나아가 구리 항상성 유지는 면역 기능을 강화하여 유방염, 간 기능 장애, 종양 발생 등 다양한 질병의 위험도를 낮출 수 있으며, 조직 염증과 섬유화의 예방을 통해 치료 비용 절감 효과도 기대할 수 있다[47,48].
장기적으로 적정 구리 관리 전략은 질병 예방과 생산성 향상을 동시에 실현하여 낙농 농가의 경제적 부담을 경감시키고, 안정적인 생산 환경 조성에 기여할 것이다. 따라서 젖소 개체의 생리적 안정성과 생산성을 뒷받침하는 구리 항상성 조절은 낙농 산업 전반에서 경제적 가치를 높이는 핵심 관리 요소로 평가될 수 있다.
결 론
본 총설에서는 젖소의 체내 구리 항상성 유지가 비유 생리와 낙농 산업에 미치는 영향을 고찰하였다. 산업화와 가축 분뇨 처리 시스템의 미비 등으로 인해 토양 및 수질 내 구리 오염 증가와 사료를 통한 과도한 구리 공급으로 인해 반추동물에서 만성 구리 중독 사례가 증가하고 있다. 특히, 구리 항상성 붕괴가 젖소의 생리적 변화에 미치는 영향에 대한 인식과 연구가 부족하여, 전 세계적으로 구리 수준 변화에 따른 젖소 건강 이상 사례가 증가하는 추세이다.
구리는 체내 다양한 효소의 보조인자로서 에너지 생성, 항산화, 면역 반응 등 다수의 생리적 과정에 필수적으로 관여한다. 그러나 구리의 과다 또는 결핍은 종 특이적 병리 현상을 유발할 수 있으며, 반추동물은 다른 종에 비해 구리 배출 능력이 제한적이고, 반추위에서 형성되는 thiomolybdates와 같은 물질로 인해 구리 흡수와 효소 활성에 영향을 받을 수 있다. 특히 젖소는 유선 발달과 우유 생산을 위해 체내 구리 수준이 정밀하게 조절되어야 하며, 구리 과다로 인한 유선 상피세포 증식 억제와 유방암 발생 등의 병리 현상이 보고된 바 있다.
이러한 병리학적 영향은 젖소의 건강과 수명을 저하시켜 우유 생산량과 품질을 감소시키며, 장기적으로 낙농 산업 전반의 생산성과 경제성에도 부정적 영향을 미친다. 따라서 젖소의 체내 구리 항상성 유지는 단순한 미량 원소 균형을 넘어, 유선 기능과 정상적인 성장, 안정적인 우유 생산, 나아가 낙농 산업의 지속 가능한 발전을 위해 필수적으로 관리되어야 한다.